
La colina es comúnmente suplementada en dietas de monogástricos porque los alimentos no logran cubrir su requerimiento nutricional.
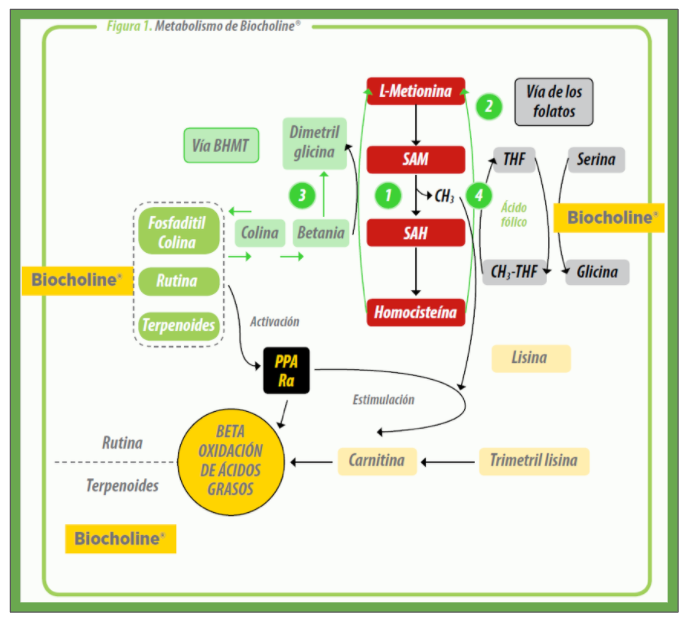
La colina cumple varias funciones metabólicas en el organismo. Como componente de la fosfatidilcolina (FC) es esencial en la estructura y funcionalidad de las membranas biológicas, también participa en el metabolismo hepático previniendo la acumulación de grasa, es un precursor del neurotransmisor acetilcolina y finalmente es un nutriente donador de grupos metilo (CH3) para el ciclo de metilación. Los ciclos de metilación en el organismo son importantes para la síntesis de ADN, creatina y fosfatidilcolina (FC).
La FC juega también un rol emulsificante y activa receptores celulares llamados PPARα (receptores proliferadores de peroxisoma), cuya activación a nivel nuclear, aumenta la oxidación de lípidos, disminuyendo su síntesis en hígado y otros tejidos. A través de su rol en el ciclo de metilación, la colina también ayuda en la regeneración de la metionina, ahorrando así este aminoácido para la síntesis de proteínas. Como tal, la suplementación con colina ayuda en última instancia al funcionamiento óptimo del metabolismo de grasas y proteínas.
En el metabolismo, la colina como tal no es donadora de grupos CH3. Para esto, primero debe ser oxidada a betaína, siendo esta última la que actúa efectivamente como donador de estas moléculas en el ciclo de metilación, permitiendo el paso de la homocisteína a metionina (Fig 1). Este rol de la betaína ha permitido su uso para reemplazar la colina suplementada en las dietas de aves. Sin embargo, la betaína no puede convertirse de vuelta a colina, por lo tanto, la betaína solo puede reemplazar a la colina en su función donadora de grupos CH3.
| La Biocholine® no solo aporta colina como FC, sino también una serie de fitoactivos como terpenoides, rutina y timol, los cuáles son potentes activadores de los PPAR-α (Fig. 1), incluso varias veces superior a la FC. Los PPARα son receptores nucleares que están relacionados con la producción de la hormona adiponectina, proteína que participa en varias funciones metabólicas favoreciendo la oxidación de ácidos grasos y la entrada de glucosa en los tejidos. |
Por lo tanto, a pesar de que tanto la Biocholine® como la betaína se utilizan para reemplazar al cloruro de colina en las dietas de aves, ambas actúan a través de mecanismos metabólicos diferentes para sustituir las funciones de la colina. Por un lado, la betaína aporta grupos CH3, que no es la función principal de Biocholine® y por otro lado, esta última cumple un rol similar a la FC en el metabolismo de los lípidos, pero en una magnitud muy superior.
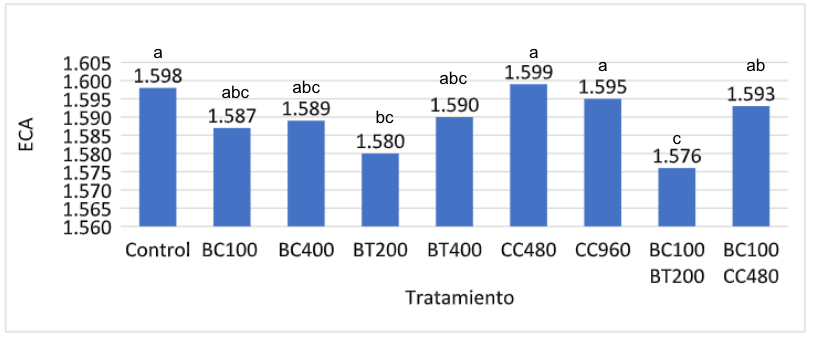
En un estudio reciente (Gonzalez et al., 2021), se observó que pollos broiler alimentados con una combinación de Biocholine® (100 g/ton) y betaína (200 g/ton) presentaron a los 42 días de edad una eficiencia de conversión alimenticia (ECA) menor (1,576) que aquellas aves alimentadas con betaína (400 g/ton) y Biocholine® (400 g/ton) en forma separada (1,590 y 1,589, respectivamente). Aun cuando esta diferencia no fue significativa (p > .05), el retorno sobre la inversión del uso combinado de betaína y Biocholine® fue 67% y 20% superior al uso de ambas por separado, respectivamente. Adicionalmente, los resultados demostraron que los pollos alimentados con la combinación de Biocholine® y betaína presentaron una ECA mas eficiente (p < .05) que las aves suplementadas con cloruro de colina.
Estos resultados abren una interrogante nutricional muy interesante respecto al rol complementario que pudieran estar ejerciendo la Biocholine® y la betaína en el metabolismo energético de las aves, ejerciendo un efecto ahorrador de la energía, lo cual debe ser investigado en mayor profundidad.

Materiales y Métodos:
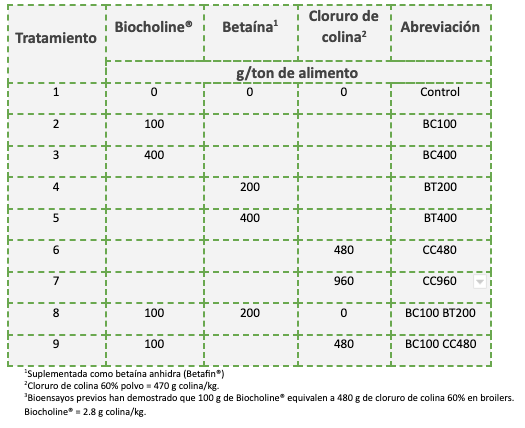
Este estudio experimental fue realizado durante el año 2020 en la estación experimental de Zootest SAS, Francia. En este, se utilizaron 450 pollos broiler machos (ROSS 308) de 1 día de edad, los cuales fueron distribuidos al azar en 9 tratamientos con 10 repeticiones cada uno (30 aves por repetición). Cada repetición consistió en un corral de 1,25 x 1,90 mt con viruta de madera nueva como material de cama. Las variables ambientales fueron controladas en forma automatizada. Los tratamientos consistieron en diferentes dosis de Biocholine®, betaína y CC (Tabla 1). El estudio tuvo una duración de 42 días.
Tabla 1: Descripción de los tratamientos experimentales:

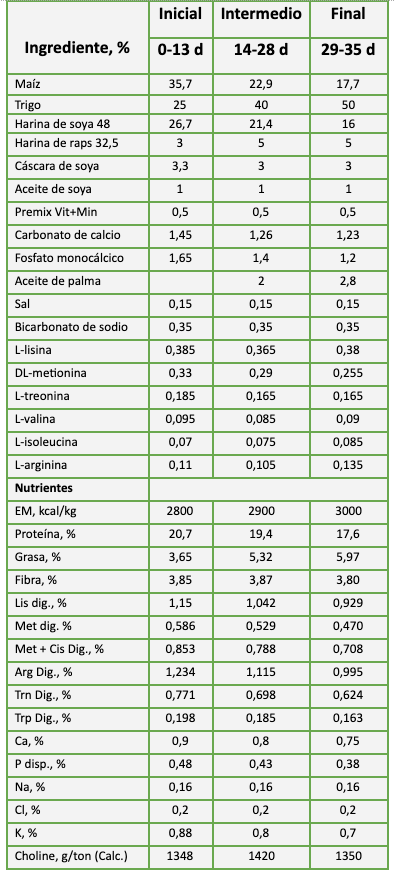
Durante el experimento, las aves fueron alimentadas con tres raciones de alimento peletizado a base de maíz, soya y trigo, inicial (0-13 d), intermedio (14-28 d) y final (29-42 d) (Tabla 2). Las principales variables medidas fueron indicadores zootécnicos y score de calidad de cama.
Tabla 2: Composición de la dieta basal:

Resultados:
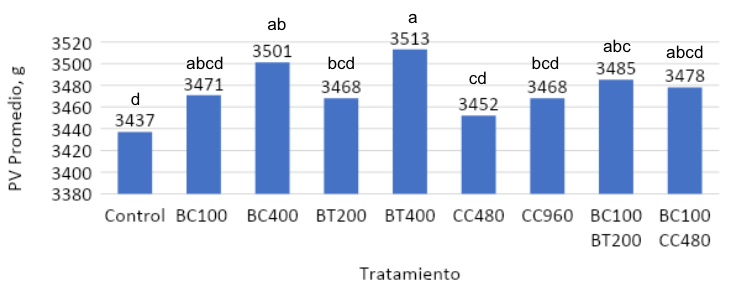
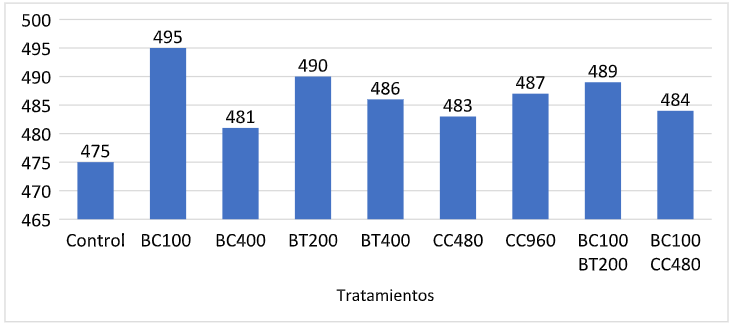
El peso vivo (PV) no presentó diferencias (p > .05) hasta los 35 d de edad. Sin embargo, a los 42 d, el mayor PV fue obtenido por los pollos alimentados con 400g/ton de betaína o Biocholine® (p >.05) (Fig. 2). Los pollos suplementados con 400g/ton de betaína presentaron un mayor PV (p <.05) que aquellos alimentados con 200 g/ton. Las aves suplementadas con 100 y 400 g/ton de Biocholine® no difirieron en PV (p > .05) de aquellos alimentados con 400 g/ton de betaína. El PV vivo de las aves alimentadas con 480 o 960 g/ton de CC 60% fue similar (p > .05) al de las aves suplementadas con 100 g/ton de Biocholine® y 200 g/ton de betaína. Los broilers alimentados con 100g/ton de Biocholine® en combinación con betaína (200 g/ton) o CC 60% (480 g/ton) obtuvieron un peso vivo intermedio que no difirió (p > .05) de los otros tratamientos (Fig. 2).
Figura 2: Resultados de peso vivo (PV) a los 42 d de edad:

La eficiencia de conversión alimenticia (ECA) no difirió (p > .05) entre los distintos tratamientos hasta los 28 d de edad. Sin embargo, tanto a los 35 como 42 d de edad se observaron diferencias (p < .05) para la ECA entre tratamientos. En la Fig. 3 se observan los resultados de ECA a los 42 d de edad.
El grupo control presentó la ECA menos eficiente, difiriendo (p <.05) de los otros tratamientos, con excepción de las aves suplementadas con CC 60%. Los pollos alimentados con la combinación de Biocholine® (100g/ton) y betaína (200g/ton), presentaron la ECA mas eficiente, sin embargo, esta no fue diferente (p > .05) de los otros tratamientos, excepto de la ECA obtenida por las aves control y aquellas alimentadas con CC 60%. No se observaron diferencias (p >.05) entre los pollos suplementados con Biocholine® y betaína, independiente de la dosis suplementada.
Figura 3: Resultados de eficiencia de conversión alimenticia (ECA) a los 42 d de edad:

Al final del estudio se calculó el Indice de Producción Europeo (IPE):
IPE = Ganancia Diaria x Sobrevivencia
10 x ECA
Aún cuando el mayor IPE se observó en los broiler suplementados con Biocholine® (100 g/ton) (Fig. 4), no se observaron diferencias (p >.05) entre tratamientos.
Figura 4: Cálculo de Indice de Producción Europeo (IPE) a los 42 d:

No se observaron diferencias (p > .05) en el score de calidad de cama entre tratamientos durante el estudio.
Conclusiones:
Los resultados zootécnicos fueron iguales o superiores a las especificaciones de la línea genética (ROSS 308), lo cual asegura la confiabilidad de los resultados.
Ambos aditivos, Biocholine® y betaína demostraron ser eficaces en el reemplazo del cloruro de colina en las dietas de pollos broiler, sin presentar diferencias significativas entre ellas.
La combinación de Biocholine® y betaína pudiesen tener un efecto positivo y sinérgico en el metabolismo energético, en especial en las aves mayores de 28 d de edad. Esta observación requiere de mayor investigación.
Autor: Javier Gonzalez, PhD, gerente técnico de Nuproxa Switzerland.
Para saber más visita: Nuproxa, efficient solutions, naturally

