



Este artículo evalúa el grado de protección de diferentes cepas vacunales de salmonella frente a cepas heterólogas mediante una amplia revisión bibliográfica.
Según el informe de la EFSA (Agencia Europea de Sanidad Alimentaria) la Salmonelosis es la segunda zoonosis en frecuencia en la Unión Europea (U.E.) después de la Campylobacteriosis con más de 94.000 casos reportados en 2016. El número de casos ha descendido significativamente desde el año 2008 hasta el 2012, manteniéndose más o menos estable desde entonces (EFSA 2017).
Salmonella spp. se compone de más de 2.500 serotipos diferentes diferenciándose según las determinantes antigénicas que presenten.
Los 4 serotipos reportados más frecuentemente en humanos en el 2016 son los responsables de aproximadamente el 71% de los casos, (ver gráfico 1).
- S.Enteritidis
- S. Typhimurium
- S. Typhimurium monofásica
- S. Infantis

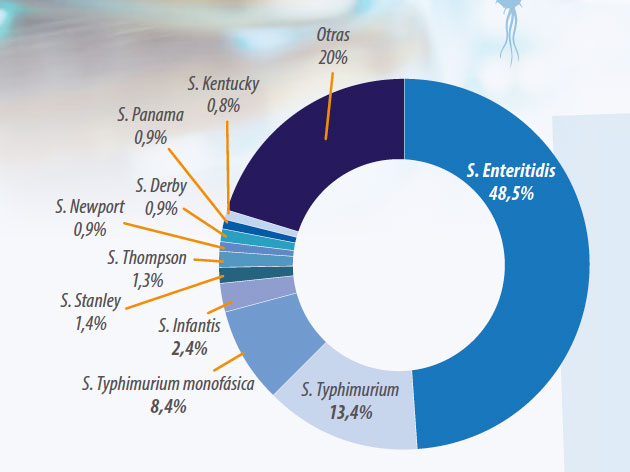
Los casos relacionados con S. Typhimurium y su variante monofásica descendieron ligeramente en 2016. Se relaciona su infección con el consumo de carne de cerdo y de rumiantes. Ocasionalmente se detectan la carne de pollo y el huevo como el origen de la infección

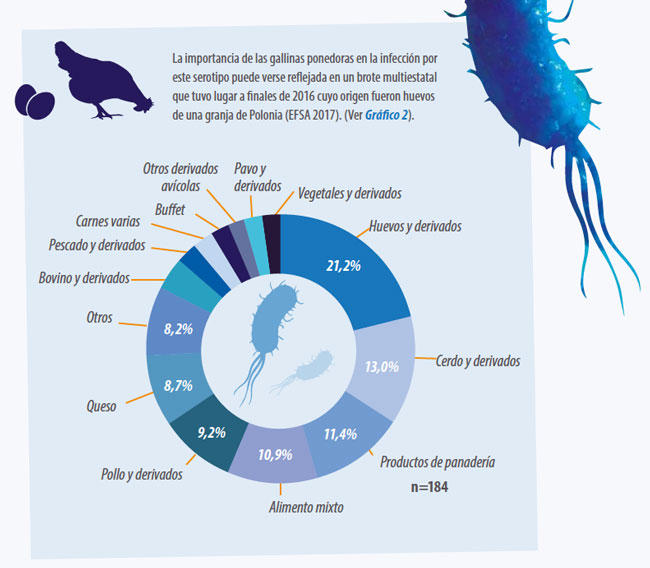

S. Infantis es el cuarto serotipo más común en humanos en Europa. Cabe remarcar que se detecta principalmente en pollos, aproximadamente el 50% de los casos positivos en pollos y derivados son de este serotipo
- En ponedoras y huevo comercial, S. Infantis es el tercer serotipo después de la S. Enteritidis y S. Typhimurium. Desafortunadamente, tal como está ocurriendo con S. Typhimurium, un número elevado de los aislamientos presenta resistencia múltiple a antibióticos (Gal-Mor et al. 2010, Franco et al. 2015, Antones et al. 2016, Hindermann et al. 2017, EFSA 2017).
A nivel de la producción primaria, los programas de control europeos monitorean los serotipos llamados de importancia zoonótica, reportándose las siguientes especies
En las manadas de reproductoras
- S. Enteritidis
- S. Typhimurium
- S. Hadar
- S. Virchow
- S. Infantis
En ponedoras
- S. Enteritidis
- S. Typhimurium
(Regulación (EC) No. 2160/2003i y Directiva 2003/99/EC).
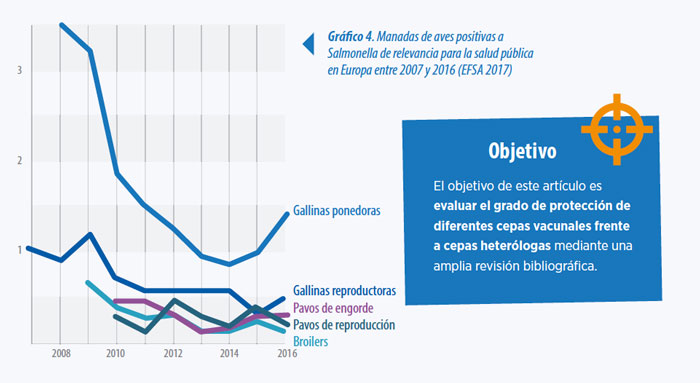
Los niveles detectados son generalmente constantes dentro de los objetivos exigidos. Sin embargo en ponedoras se observó un incremento significativo en la prevalencia de S. Enteritidis en 2015 y 2016 (Ver Gráfico 3 y Gráfico 4).

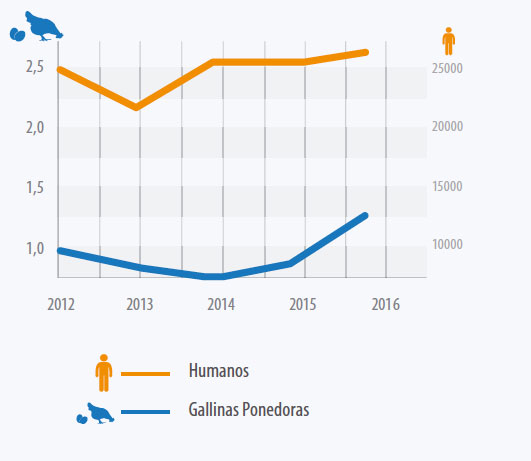
Gráfico 3. Porcentaje de manadas de gallinas ponedoras positivas a S. Enteritidis y número de casos en humanos en la U.E entre 2012 y 2016 (EFSA 2017)

La inmunización con cepas vivas
La vacunación se considera una herramienta principal en los programas de control, usándose como parte de los programas de prevención de Salmonella en avicultura desde el 1994, primero en reproductoras y más tarde en ponedoras. La marcada reducción en la incidencia de Salmonella en la última década es una prueba del éxito de los programas de prevención basándose en estrictas pautas de bioseguridad y en la profilaxis vacunal (O’Brien 2013, Lane et al. 2014). Es generalmente aceptado que la eficacia de las vacunas vivas contra la infección con cepas de campo es superior a la de las vacunas inactivadas (Revolledo y Ferreira 2012, Barrow P.A 1997).
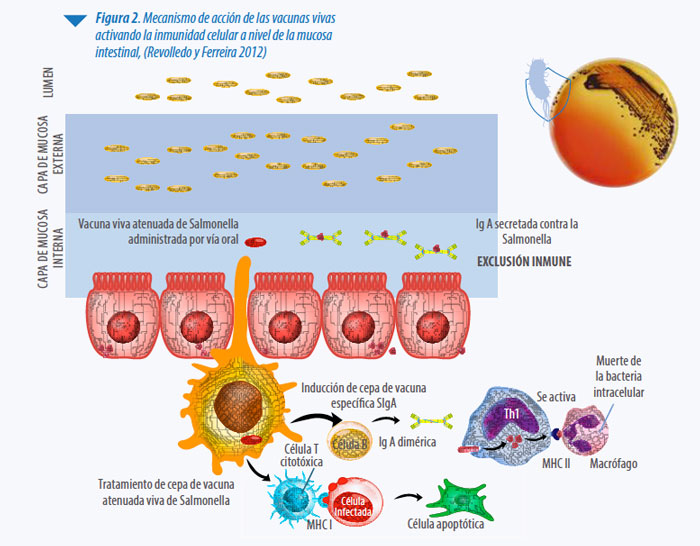
El mecanismo de acción de las cepas vacunales vivas está ampliamente documentado, ejerciendo su acción imitando las vías de infección naturales de la bacteria, estimulando ambas ramas del sistema inmune: la humoral, vía la estimulación de la excreción de IgA a nivel de la mucosa intestinal, y la celular activando la respuesta inespecífica al ser fagocitada por macrófagos, procesada y presentada a las células T (ver Figura 2).
Las cepas vivas presentan un marcado fenómeno de exclusión competitiva, cuyo efecto se obtiene a pocas horas de la aplicación de la cepa viva (Van Immerseel et al. 2005, Methner et al. 1997,2010 y 2011). En cambio, las vacunas inactivadas estimulan principalmente la rama humoral del sistema inmune (producción de anticuerpos). Al ser la Salmonella una bacteria intracelular, la inmunidad mediada por células ejerce un papel principal en su control (Revolledo y Ferreira 2012, Barrow P.A 1997).

Protección homóloga frente a heteróloga
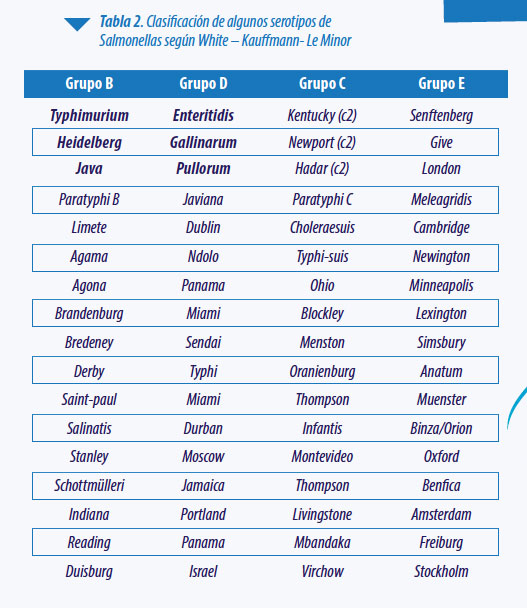
La protección conferida por las vacunas es, principalmente, serotipo específica. Numerosos estudios demuestran que la protección homóloga (conferida por una vacuna de un serotipo similar a la cepa de desafío) es superior a la heteróloga (conferida por una vacuna de un serotipo diferente a la cepa de desafío). Sin embargo, esta también demostrado que la protección cruzada entre serotipos dentro del mismo grupo (según la clasificación White – Kauffmann- Le Minor) es superior a la protección cruzada frente a cepas que no pertenecen al mismo.
Los serotipos más destacables dentro del serogrupo “B” son:
- S. Typhimurium
- S. Heidelberg
- S. Java
Los serotipos más relevantes dentro del serogrupo “D” son:
- S. Enteritidis
- S. Gallinarum
- S. Pullorum
(Barrow y Methner 2013)

Vacunación con S. Typhimurium
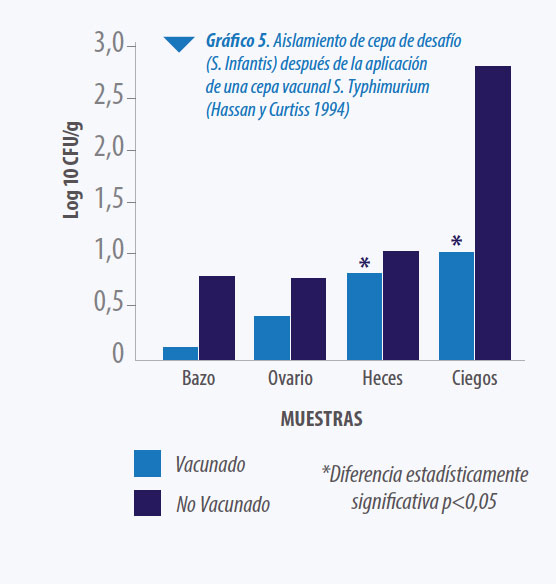
Varios autores estudiaron la respuesta de la protección conferida por la vacunación con un serotipo especifico frente al desafío con serotipos diferentes. Ya en 1994 Hassan y Curtiss estudiaron el efecto de la vacunación con S. Typhimurium cepa x3985 frente al desafío con diferentes serotipos, y pudieron demostrar una protección excelente frente a otros serotipos del mismo grupo B. Asimismo, resultó alta la eficacia frente a Salmonellas del grupo E y D y relativamente alta frente a grupo C.
Resulta interesante la observación que la protección frente a la infección del ovario fue excelente tanto cuando el desafío se realizó con S. Typhimurium como con S.Enteritidis. (Ver Gráfico 5)

Lee ha demostrado que la aplicación de una cepa atenuada de S. Typhimurium (cepa JOL912) produjo una protección eficaz frente al desafío con diferentes cepas del grupo D (S. Enteritidis y S. Gallinarum) (Lee 2015). De hecho, la vacuna viva de S. Typhimurium cepa Nal 2/Rif 9/Rtt (componente de la vacuna comercial AviPro™ Salmonella DUO) fue la primera en comercializarse en Europa. En Israel, donde la prevalencia de S. Enteritidis era alta, lograron controlar este serotipo usando la vacuna de S. Typhimurium (Elkin N.comm. pers.). Una experiencia similar se obtuvo en Alemania (Lohren U .comm, pers.).
Protección cruzada entre S. Typhimurium y S. Typhimurium monofásica
La mayoría de las cepas de Salmonella son móviles mediante flagelos, estos están codificados por un par de genes (fljB, fljC), cuya expresión está regulada por un mecanismo denominado “Variación de Fase”.
En las cepas de S. Typhimurium el antígeno H tiene 2 fases:
- Una llamada “i”
- La segunda “1,2”
Las cepas que demuestran estructura antigénica similar a la de S. Typhimurium, pero sin expresar ciertos antígenos flagelares, se denominan ‘Salmonella Typhimurium like strains’.
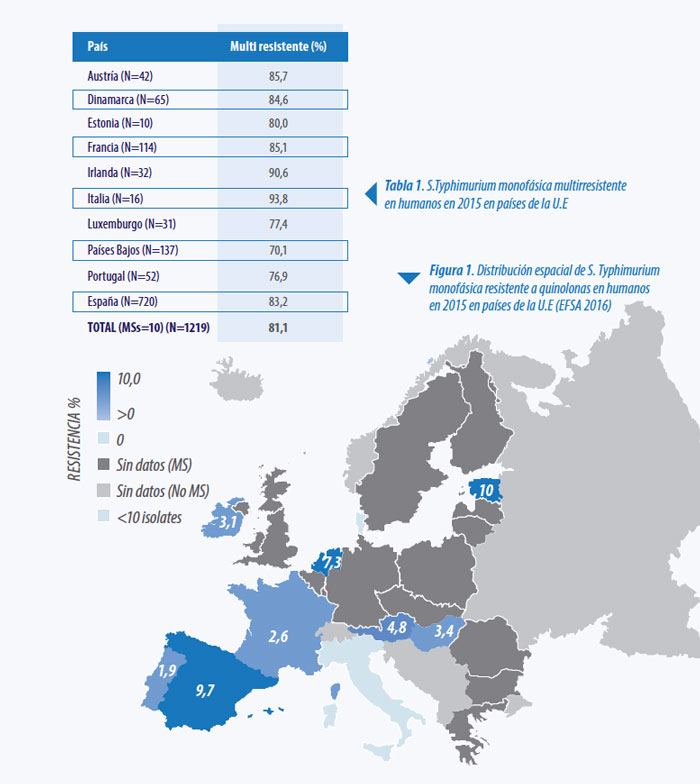
Desde mediados de los 90 se han publicado numerosos informes en varios países europeos, así como en EE.UU. y otros países, sobre el aislamiento de cepas de ‘Salmonella Typhimurium like strains’ (EFSA Journal 2010, Andrea et al. 2008). El serotipo S. Typhimurium 1,4,[5],12:i:- y el S. Typhimurium 1,4,[5],12:i:1,2 tienen las mismas determinantes antigénicas O y Hi. En la cepa 1,4,[5],12:i:- falta la fase 2 flagelar y por ello se denomina “S. Typhimurium monofásica”, siendo actualmente el tercero en frecuencia en la infecciones detectadas en humanos en Europa y presentando muchas cepas resitencias multiples a antibióticos (EFSA 2017)
.Kilroy y colaboradores evaluaron en 2015 la eficacia de la vacunación con S. Typhimurium cepa Nal 2/Rif 9/Rtt contra la infección con S. Typhimurium monofásica. Se pudo demostrar que la excreción de la cepa de desafío fue reducida significativamente en las aves vacunadas en comparación con las aves no vacunadas. Así también se pudo demostrar que la presencia de la cepa de exposición en el bazo y ciegos fue reducida en las aves vacunadas (Ver Gráfico 6) (Kilroy et al.2015).
La Vacunación con S. Typhimurium en combinación con S. Enteritidis
Actualmente se comercializan vacunas vivas frente a los serotipos S. Enteritidis y S. Typhimurium solamente. La vacunación con S. Enteritidis y S. Typhimurium exhibe un cierto grado de protección cruzada contra serotipos diferentes, ya que la protección no solamente depende de la clasificación del serotipo sino tambien de la composición genetica completa de cada cepa (Varmuzova et al. 2016). Algunos investigadores plantearon la pregunta si la inclusión de varios serotipos podría potenciar la respuesta homóloga. Gantois y colaboradores evaluaron la eficacia de la vacunación con una vacuna compuesta por 2 cepas en un modelo de infección intravenosa con cepa virulenta de S. Enteritidis: S. Enteritidis cepa Sm24/Rif12/Ssq S. Typhimurium cepa Nal 2/Rif 9/Rtt

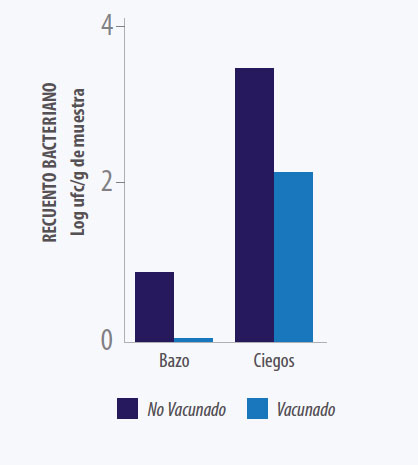
Gráfico 6. Determinación cuantitativa del número de bacterias
monofásicas (S. enterica subsp. Enterica Serovar 1,4,[5],12:i:– )
en ciego y bazo 14 días post-infección. (Kilroy et al. 2015).

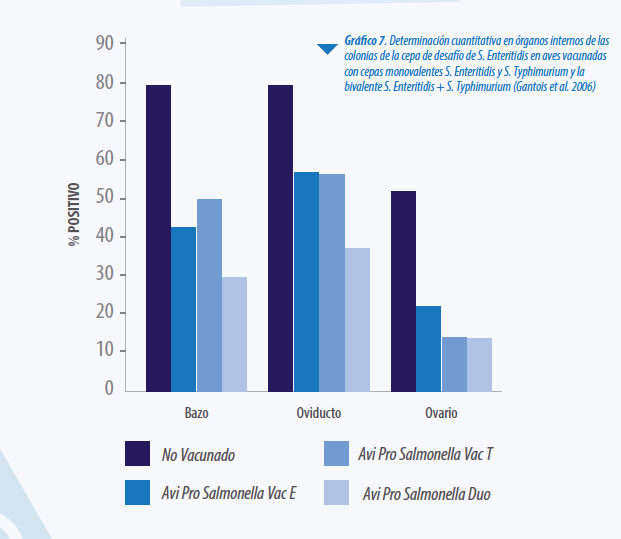
En otro estudio realizado por Matulova y colaboradores en 2013 se vacunaron aves con ambas cepas S.Enteritidis y S. Typhimurium (Cepas SPI1), demostrándose, en este caso también, mayor protección comparado con la vacunación con cada cepa por separado.
Se puede concluir que la aplicación conjunta de más de una cepa tiene un efecto “sinérgico”, aumentando la eficacia frente a ambas cepas incluidas en la vacuna (Matulova et al. 2013)
Vacunación frente a S. Infantis
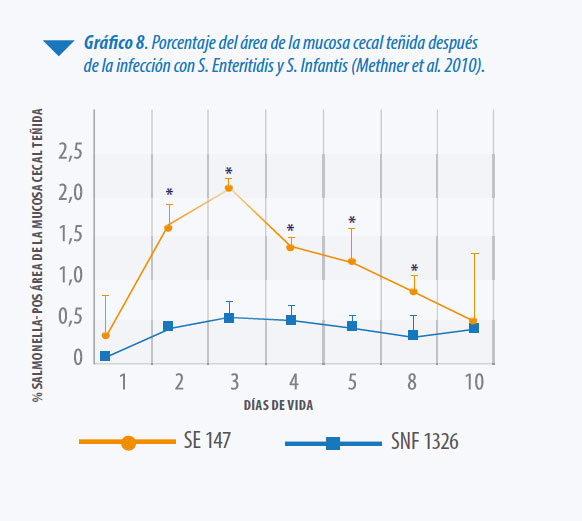
Comparando con otros serotipos como S. Enteritidis y S. Typhimurium, S. Infantis, tiene una mayor habilidad de colonizar los ciegos, pero su poder de invasión del organismo es relativamente limitado (Methner et al. 2010, Berndt et al 2007) (Ver Gráfico 8). La protección a nivel del tracto intestinal cobra el serotipo S. Infantis resulta de aún mayor importancia (Berndt et al. 2007, Setta et al. 2012). Tal como fue mencionado, las vacunas vivas proporcionan una protección marcada a nivel del sistema inmune asociado a mucosas, con demostrado efecto sobre la producción de IgA (Revolledo 2012).
Al no existir vacunas vivas frente al serotipo S. Infantis, varios investigadores evaluaron el efecto de la vacunación con serotipos diferentes, asumiendo que el efecto de la exclusión competitiva, así como, cierto grado de reacción cruzada, podría conferir mayor o menor grado de protección. Al vacunar aves el primer día de edad con S. Enteritidis e infectar con diferentes cepas de campo: S. Enteritidis, S. Infantis y S. Typhimurium, se pudo observar que S. Enteritidis indujo una reacción marcada a nivel de la mucosa cecal inhibiendo la colonización, tanto de cepas homólogas como heterólogas administradas 24 horas después, mientras que la vacunación con S. Infantis solo pudo inducir inhibición de la colonización de cepas homólogas (Methner et al. 2010).


- La prevalencia de S. Enteritidis en gallinas ponedoras aumentó en 2015 y 2016 después de mostrar una tendencia marcada a la baja en la última década (EFSA 2017).
- Los casos relacionados con S. Typhimurium y su variante monofásica se relacionan con cerdos y rumiantes principalmente, aunque son frecuentes también en la avicultura, tal vez relacionada su detección con granjas avícolas en cercanías de las porcinas.
- S. Infantis se detecta principalmente en pollos.
- Las vacunas vivas de Salmonella constituyen un pilar importante en la prevención de salmonelosis en las manadas de reproductoras y ponedoras.
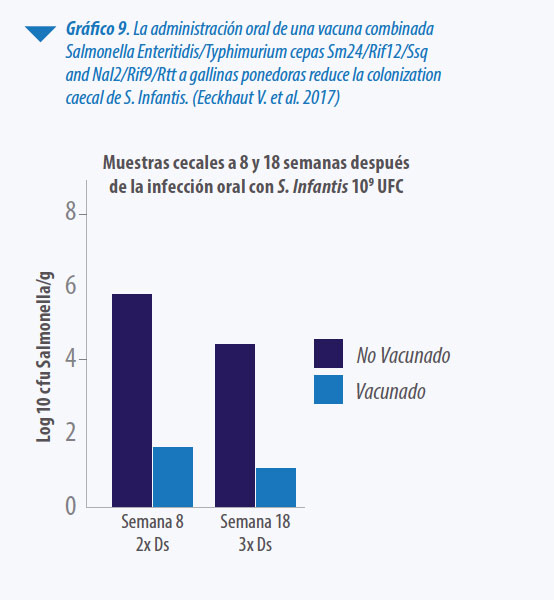
- El efecto de las vacunas vivas radica tanto en el fenómeno de la exclusión competitiva como en la estimulación de la inmunidad celular y es, principalmente, serotipo específico. Sin embargo, en el caso particular de una vacuna bivalente de S. Enteritidis y S. Typhimurium, estudios recientes demuestran niveles de protección cruzada importante frente a un serotipo del grupo C (Eeckhaut et al. 2017).
- Considerando lo anterior, en el caso de infecciones contra las cuales no existan vacunas vivas del serotipo específico de Salmonella, se recomienda emplear vacunas vivas heterólogas, ya que puedan ejercer una protección derivada del efecto de la exclusión competitiva, así como sensibilizar el sistema inmune inespecífico.
- Se pudo demostrar que la combinación de ambas cepas Enteritidis y Typhimurium potencia la protección tanto frente a cepas homólogas como heterólogas (Gantois 2006, Methner et al 2011, Matulova et al. 2013).
- Se podria potenciar la respuesta específica usando además de las vacunas vivas con cepas heterólogas, vacunas inactivadas homólogas (Penha et al. 2012, Dorea et al. 2010, Deguchi et al. 2009, Bailey et al. 2007). En el caso de que no existiera una vacuna inactivada comercializada contra el serotipo homólogo, se podría considerar el uso de autovacunas inactivadas (Pavic 2010, Bailey et al. 20072).
ESBRLSDU00002

