



La enfermedad de Newcastle, ENC, es considerada una de las enfermedades infecciosas más importantes de las aves, porque las cepas velogénicas del virus pueden causar brotes con alta morbilidad, mortalidad y restricción del comercio internacional, ocasionando importantes pérdidas económicas a la industria avícola, por ello, la enfermedad forma parte de la lista de enfermedades notificables a la Organización Mundial de Salud Animal (Miller y Koch, 2020).
La enfermedad de Newcastle se previene con bioseguridad y vacunación, y aun cuando varios tipos de vacunas efectivas, vivas e inactivadas son aplicadas, la ENC continúa siendo un problema en muchos países del mundo.
EL AGENTE CAUSAL
La base de datos del comité internacional de taxonomía viral clasifica al virus como familia Paramyxoviridae, subfamilia Avulavirinae, esta última distribuida en tres géneros:
- Orthoavulavirus.
- Metaavulavirus.
- Paravulavirus.
Los paramixovirus aviares han sido aislados de diferentes especies de aves, y son clasificados en 21 serotipos por pruebas serológicas y análisis filogenético (WOAH, 2021).
La enfermedad de Newcastle es causada por cepas virulentas de Paramixovirus aviar tipo 1 (APMV-1), especie Orthoavulavirus aviar tipo -1 (OAV-1).
El virus de la enfermedad de Newcastle (VEN), tiene un genoma de ARN de cadena simple, no segmentado, de sentido negativo, que mide 15,186 nucleótidos (Alexander, 2003). El virión presenta una envoltura cubierta con dos capas lipídicas derivada de la membrana plasmática de la célula hospedera (Mast y Demeestere, 2009).
El genoma del virus está compuesto por seis genes en orden 3´-NP-P-M-F-HN-L-5´, que codifican siete proteínas virales:
- Nucleoproteína (NP),
- Fosfoproteína (P),
- Proteína de la matriz (M),
- Proteína de fusión (F),
- Hemaglutinina-neuraminidasa (HN) y
- RNA polimerasa, llamada la polimerasa grande (L);
La edición del ARN de la proteína P produce una proteína adicional, la proteína V, con actividad anti-interferón, que permite al virus contrarrestar la respuesta innata de la célula huésped (Miller y Koch, 2020) (Figura 1).

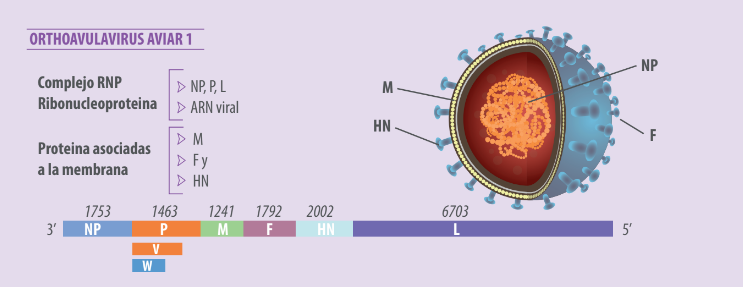
Figura 1. Representación esquemática del genoma del virus de la ENC y sus proteínas.


La principal propiedad biológica del virus es aglutinar glóbulos rojos de aves, anfibios y reptiles, esto debido a la acción de la proteína HN sobre los receptores de ácido siálico de la superficie de los glóbulos rojos.
- La hemoaglutinación (HA) viral permite determinar la presencia viral en cultivos virales y fluido alantoideo de embriones de pollo (Miller y Koch, 2020), y cuantificar los anticuerpos en el suero de las aves mediante la prueba de inhibición de la hemoaglutinación (IHA).
Desde hace algún tiempo viene siendo investigado el efecto oncolítico de algunas cepas del VEN sobre las células tumorales humanas y su uso como tratamiento contra el cáncer en humanos, este efecto está asociado a la actividad anti-interferón tipo I de la proteína V, la selectiva replicación que tiene el virus en células tumorales se debe a defectos de estas en la activación de las vías de señalización del IFN tipo I y en las vías apoptóticas entre otras (Schirrmacher, 2017).
REPLICACIÓN VIRAL
El VEN se replica en el citoplasma de la célula. La hemaglutinina (HN) reconoce al receptor celular, activando a la proteína F a fusionar las membranas viral y celular, permitiendo la entrada viral al citoplasma por endocitosis (Bergfeld, 2017; Miller y Koch, 2020).
- El complejo ribonucleoproteina (RPN) contiene el genoma de ARN envuelto con la nucleoproteína (NP) asociado al complejo polimerasa compuesto por la fosfoproteína (P) y la proteína grande (L);
- Después de la entrada de la nucleocapside viral al citoplasma, esta se disocia de la proteína M y se libera para iniciar la síntesis del ARNm requerido para la traducción de las proteínas virales.
- La proteína P media la unión del complejo de polimerasa a la nucleocapside y la proteína L realiza las actividades catalíticas de la polimerasa (Dortmans et al., 2011; Cox y Plemper, 2017).

Todos los genes codifican para una sola proteína principal, sin embargo, el ARNm del gen P, resulta en la formación de dos proteínas no estructurales la proteína V y la proteína W (Vilela et al., 2022).
- Después que el ARNm es traducido en proteínas virales, el genoma de polaridad negativa se replica, produciendo un ARN anti-genómico que sirve como molde para la síntesis de un ARN genómico de longitud completa (Miller y Koch, 2020; Dortmans et al., 2011).
- Las proteínas HN sintetizadas en la célula son transportadas a la membrana celular insertándose en ella, a ello sigue el alineamiento de la nucleocapside y el ARN viral a las regiones modificadas cercanas de la membrana celular, que contienen las glicoproteínas virales (Miller y Koch, 2020);
- La proteína M de la matriz, organiza el ensamblaje de las partículas virales a través de la interacción con las proteínas N del complejo RNP y los complejos glicoproteicos integrados a la membrana (Cox y Plemper, 2017).
- Las nuevas partículas virales salen por gemación brotando de la superficie celular, arrastrando la envoltura lipoproteica de la célula (Miller y Koch, 2020; Cox y Plemper, 2017).
- La neuraminidasa de la proteína HN permite liberar al virus de la célula, removiendo el receptor celular (Bergfeld, 2017).
PATOGENICIDAD VIRAL
De acuerdo con los signos clínicos que ocasionan en las aves infectadas, las cepas del VEN se clasifican en:
- Cepas velogénicas viscerotrópicas (vvVEN) y cepas mesogénicas, (ambas causantes de la enfermedad de Newcastle) y,
- Cepas lentogénicas y entéricas o apatógenas (usadas como vacunas).
La Organización Mundial de Salud Animal, OMSA, ha determinado que una cepa de APMV-1 es virulenta, cuando tienen:
- Un índice de patogenicidad intracraneana (IPIC) igual o mayor a 0,7 (teniendo en consideración los valores mínimos y máximos de 0 a 2) o,
- El sitio de escisión de la proteína F es de múltiples aminoácidos básicos y tiene una fenilalanina en la posición 117 (WOAH, 2021).
La glicoproteína F es la clave de virulencia y patogénesis viral, debido a que el ingreso a la célula depende de la fusión de la membrana celular y viral, después de la escisión de la proteína F0 en F1 y F2, por las proteasas del huésped (Miller y Koch, 2020).
La diferencia entre un virus virulento y uno no virulento es que:
- Los virus con una secuencia de aminoácidos monobásica en el sitio de escisión de F0 son apatógenos porque:
- Este substrato es susceptible solo a enzimas excretadas en mucosas, como tripsina, por lo tanto, ocasionan una infección localizada.
- Contrariamente los virus con una secuencia dibásica son patógenos porque son:
- Susceptibles de enzimas intracelulares ubicuas, que permiten al virus diseminarse y causar una infección sistémica y mortal (Czeglédi et al., 2006; Miller y Koch, 2020).
ÍNDICE DE PATOGENICIDAD INTRACRANEANA
El índice de patogenicidad intracraneana (IPIC), es el mejor método para determinar patogenicidad viral porque no solo determina la patogenicidad de las cepas, sino que cuantifica en valores.
- El fluido alantoideo confirmado positivo por HA con un título hemaglutinante mayor a 1:16 es diluido 1/10 en solución salina e inyectado intracranealmente a una dosis de 0,05 mL a diez pollos SPF de 1 día de edad.
- Las aves son examinadas diariamente durante 8 días, asignándoles un score diario de: 0 si normales, 1 si enfermas y 2 si mueren.
- Las aves muertas son registradas como 2 en cada uno de los días siguientes hasta los 8 días.
- El IPIC se calcula con el promedio de score diario por ave durante los ocho días; los virus más virulentos se acercan al score máximo de 2,0 y los virus lentogénicos a 0,0.
Se considera que un virus es patógeno cuando tiene un IPIC igual o mayor a 0,7 (WOAH, 2021). Existen grandes diferencias en patogenicidad entre cepas del virus. El cuadro 1, resume el índice de patogenicidad de los diferentes patotipos usando diferentes pruebas (Alexander, 1989 y 1998).

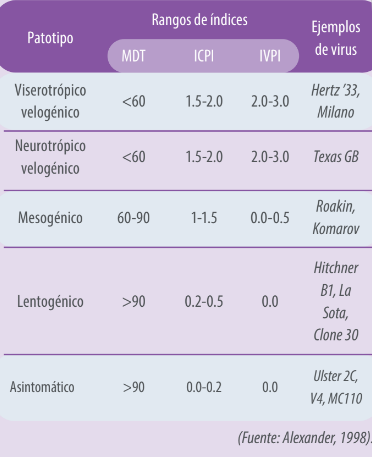
Cuadro 1. Patotipos e índice de patogenicidad del virus de la ENC.
Se considera que todos los aislamientos de APMV-1 pertenecen a un solo serotipo, sin embargo, algunas variaciones antigénicas han sido demostradas entre diferentes aislamientos mediante diversas pruebas y, aun cuando no existen variaciones antigénicas entre cepas, el análisis genético se ha convertido en el principal método de caracterización y ha reemplazado al uso de mAbs para tipificar los aislamientos de los virus de la enfermedad de Newcastle.
DETERMINACIÓN DE GENOTIPOS VIRALES
Los virus de la Enfermedad de Newcastle tienen bajas tasas de recombinación, sin embargo, a través del tiempo se han detectado ciertas diferencias antigénicas que han motivado la clasificación de las cepas dentro de linajes o genotipos. De acuerdo con esto las cepas son agrupadas en dos principales clases, virus de clase I y virus de clase II;
- Los virus de clase I son de baja patogenicidad y son encontrados en aves silvestres,
- Mientras que los de clase II, están clasificados en múltiples genotipos y pueden ser no patógenos o virulentos (Cuadro 2.).
La caracterización del genotipo del VEN, se logra secuenciando el gen F completo, lo que además permite determinar su virulencia;
- Para identificar nuevos genotipos es usado un sistema basado en una distancia evolutiva media de 10% entre grupos genéticos (Dimitrov KM et al 2016; Dimitrov KM et al, 2019).
El desarrollo de secuenciación de próxima generación, en inglés Next-generation sequencing (NGS), ha surgido como herramienta para investigación y caracterización genética de patógenos.
- Sin embargo, este método es ideal para los virus RNA que mutan rápidamente, donde se requiere además analizar aspectos evolutivos (Butt et al., 2019).

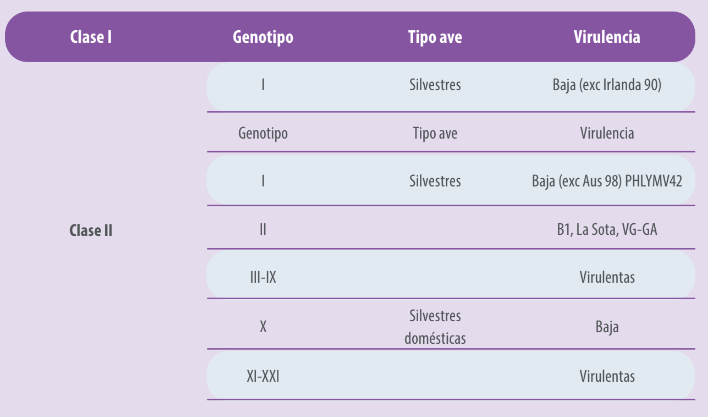
Cuadro 2. Clasificación de genotipos del virus.
La clasificación actual de genotipos incluye tres nuevos genotipos (XIX, XX y XXI), haciendo un total de 21 genotipos virales dentro de los virus de clase II (Dimitrov et al. (2019).
RESISTENCIA A LOS AGENTES FÍSICOS Y QUÍMICOS
Por ser un virus con envoltura, la viabilidad de los Paramixovirus aviares es destruida fácilmente por agentes físicos y químicos tales como:
- Calor,
- Luz ultravioleta,
- Procesos de oxidación,
- pH y
- Tratamiento con la mayoría de los desinfectantes.
No obstante, las publicaciones referentes a tiempos de supervivencia del virus fuera del huésped muestran diferencias atribuidas a temperatura, humedad ambiental y el medio en el cual el virus fue analizado (Kinde et al., 2004).

