



Konten ini tersedia dalam: English ไทย (Thai) Tiếng Việt (Vietnamese)
Newcastle disease (ND) dianggap sebagai salah satu penyakit menular yang paling penting pada unggas karena strain virus yang bersifat velogenik dapat menyebabkan wabah dengan morbiditas dan mortalitas yang tinggi dan pembatasan perdagangan internasional.
- Hal ini menyebabkan kerugian ekonomi yang signifikan pada industri perunggasan. Inilah sebabnya mengapa penyakit ini termasuk dalam daftar penyakit yang harus dilaporkan ke Organisasi Kesehatan Hewan Dunia (Miller dan Koch, 2020).
- Penyakit Newcastle dicegah dengan biosekuriti dan vaksinasi, dan meskipun beberapa jenis vaksin hidup dan vaksin tidak aktif yang efektif telah digunakan, ND terus menjadi masalah di banyak negara di seluruh dunia.
AGEN PENYEBAB
Basis data Komite Internasional untuk Taksonomi Virus mengklasifikasikan virus ini sebagai famili Paramyxoviridae, subfamili Avulavirinae, yang didistribusikan ke dalam tiga genera:
- Orthoavulavirus.
- Metaavulavirus.
- Paravulavirus.
Avian paramyxovirus telah diisolasi dari berbagai spesies unggas, dan diklasifikasikan ke dalam 21 serotipe melalui tes serologis dan analisis filogenetik (WOAH, 2021).
- Penyakit Newcastle disebabkan oleh strain virulen Avian Paramyxovirus tipe 1 (APMV-1), spesies Avian Orthoavulavirus tipe -1 (OAV-1).
Newcastle disease virus (NDV) memiliki genom RNA untai tunggal, tidak tersegmentasi, dan berasa negatif dengan ukuran 15.186 nukleotida (Alexander, 2003).
Virion memiliki lipid bilayer amplop yang berasal dari membran plasma dari sel inang (Mast y Demeestere, 2009).
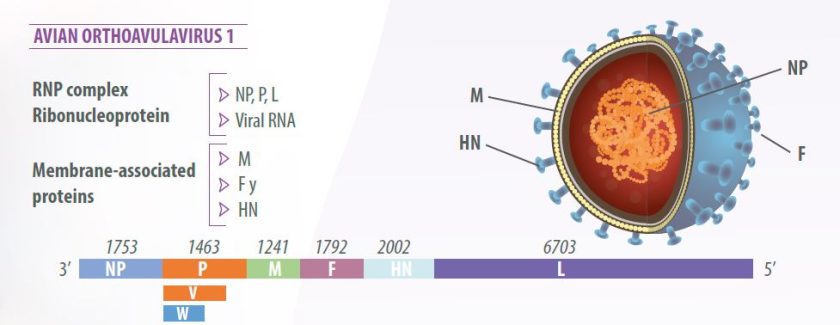
Genom virus ini terdiri dari enam gen dalam urutan 3′-NP-P-M-FHN- L-5′, yang mengkode tujuh protein virus:
- Nucleoprotein (NP),
- Phosphoprotein (P),
- Matrix protein (M),
- Fusion protein (F),
- Hemaglutinin-neuraminidase (HN) dan
- RNA polimerase, disebut polimerase besar (L).
Pengeditan RNA dari protein P menghasilkan protein tambahan, protein V, dengan aktivitas antiinterferon, yang memungkinkan virus untuk melawan respons sel inang bawaan (Miller dan Koch, 2020) (Gambar 1).

Sifat biologis utama virus ini adalah menggumpalkan sel darah merah burung, amfibi, dan reptil, karena aksi protein HN pada reseptor asam sialat pada permukaan sel darah merah.
Hemaglutinasi virus (HA) memungkinkan penentuan keberadaan virus dalam kultur virus dan cairan alantoik dari embrio ayam (Miller dan Koch, 2020), dan kuantifikasi antibodi dalam serum unggas dengan uji penghambatan hemaglutinasi (HAI).
Efek onkolitik dari beberapa jenis NDV pada sel tumor manusia dan penggunaannya sebagai pengobatan kanker pada manusia telah diteliti selama beberapa waktu.
- Efek ini dikaitkan dengan aktivitas anti-interferon tipe I protein V.
Replikasi selektif yang dimiliki virus dalam sel tumor disebabkan oleh cacatnya sel-sel ini dalam aktivasi jalur pensinyalan IFN tipe I dan jalur apoptosis antara lain (Schirrmacher, 2017).
REPLIKASI VIRUS
NDV bereplikasi di dalam sitoplasma sel. Hemaglutinin (HN) mengenali reseptor seluler, mengaktifkan protein F untuk menyatukan virus dan membran sel, memungkinkan masuknya virus ke dalam sitoplasma melalui endositosis (Bergfeld, 2017; Miller dan Koch, 2020).
- Kompleks ribonukleoprotein (RPN) berisi genom RNA yang dibungkus dengan nukleoprotein (NP) yang terkait dengan kompleks polimerase yang terdiri dari fosfoprotein (P) dan protein besar (L);
- Setelah nukleokapsida virus masuk ke dalam sitoplasma, nukleokapsida virus akan terlepas dari protein M dan dilepaskan untuk memulai sintesis mRNA yang diperlukan untuk penerjemahan protein virus.
- Protein P memediasi pengikatan kompleks polimerase ke nukleokapsida dan protein L melakukan aktivitas katalitik polimerase (Dortmans et al., 2011; Cox dan Plemper, 2017).
- Semua gen mengkode satu protein utama, namun MRNA gen P menghasilkan pembentukan dua protein non-struktural, protein V dan protein W (Vilela et al., 2022).

(1) After the mRNA is translated into viral proteins, the negative polarity genome replicates, producing an anti-genomic RNA that serves as a template for the synthesis of a full-length genomic RNA (Miller and Koch, 2020; Dortmans et al., 2011).
(2) HN proteins synthesized in the cell are transported to the cell membrane by insertion, followed by the alignment of the nucleocapside and viral RNA to the nearby modi ed regions of the cell membrane, which contain the viral glycoproteins (Miller and Koch, 2020);
- Matrix protein M, organizes the assembly of viral particles through interaction with N proteins of the RNP complex and membrane-integrated glycoprotein complexes (Cox and Plemper, 2017).
(3) New viral particles bud out by budding from the cell surface, dragging the lipoprotein envelope of the cell (Miller and Koch, 2020; Cox and Plemper, 2017).
(4) The neuraminidase of the HN protein allows the virus to be released from the cell by removing the cellular receptor (Bergfeld, 2017).
PATOGENISITAS VIRUS
Berdasarkan tanda-tanda klinis yang ditimbulkannya pada unggas yang terinfeksi, Strain NDV diklasifikasikan sebagai berikut:
- Strain viscerotropik velogenik (vvVEN) dan strain mesogenik (keduanya menyebabkan Newcastle disease) dan,
- Strain lentogenik dan enterik atau apatogenik (digunakan sebagai vaksin).
Organisasi Dunia untuk Kesehatan Hewan, WOAH, telah menetapkan bahwa suatu strain APMV-1 bersifat virulen jika memiliki:
- Indeks patogenisitas intrakranial (IPIC) sama dengan atau lebih besar dari 0,7 (dengan mempertimbangkan nilai minimum dan maksimum dari 0 hingga 2) atau,
- Situs pembelahan protein-F adalah beberapa asam amino basa dan memiliki fenilalanin pada posisi 117 (WOAH, 2021).
F-glikoprotein adalah kunci virulensi dan patogenesis virus , karena masuknya virus ke dalam sel bergantung pada fusi virus dan membran sel setelah pembelahan protein F0 menjadi F1 dan F2 oleh protease inang (Miller dan Koch, 2020).
Perbedaan antara virus yang ganas dan virus yang tidak ganas adalah:
- Virus dengan urutan asam amino monobasa pada situs pembelahan F0 bersifat apatogenik karena: Substrat ini hanya rentan terhadap enzim yang diekskresikan dalam selaput lendir, seperti tripsin, sehingga menyebabkan infeksi lokal.
- Sebaliknya, virus dengan urutan dibasa bersifat patogen karena: Rentan terhadap enzim intraseluler yang ada di mana-mana, yang memungkinkan virus menyebar dan menyebabkan infeksi sistemik dan fatal (Czeglédi et al., 2006; Miller dan Koch, 2020).
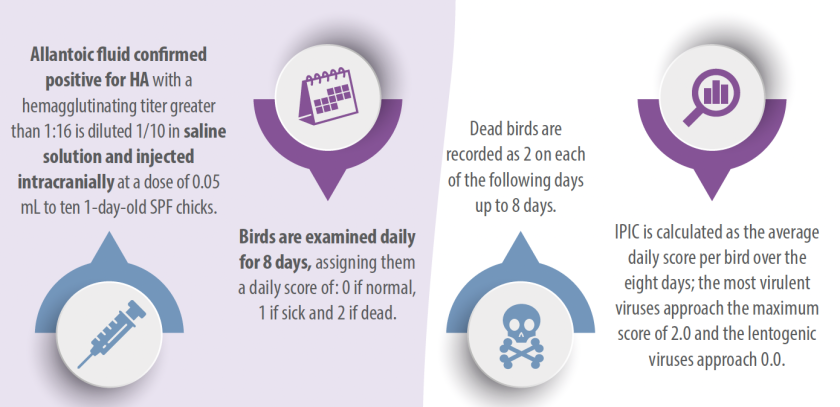
INDEKS PATOGENISITAS INTRAKRANIAL
Indeks patogenisitas intrakranial (IPIC) adalah metode terbaik untuk menentukan patogenisitas virus karena tidak hanya menentukan patogenisitas galur, tetapi juga mengukur nilainya.
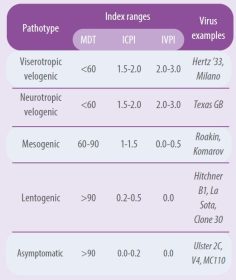
Virus dianggap patogen jika memiliki IPIC sama dengan atau lebih besar dari 0,7 (WOAH, 2021). Ada perbedaan besar dalam patogenisitas di antara strain virus. Tabel 1 merangkum indeks patogenisitas dari berbagai patotipe dengan menggunakan tes yang berbeda (Alexander, 1989 dan 1998).

Tabel 1. Patotipe dan indeks patogenisitas virus ND

Semua isolat APMV-1 dianggap termasuk dalam satu serotipenamun, beberapa variasi antigenik telah ditunjukkan di antara isolat yang berbeda dengan berbagai pengujian dan, bahkan ketika tidak ada variasi antigenik di antara galur, analisis genetik telah menjadi metode utama untuk karakterisasi dan telah menggantikan penggunaan mAbs untuk mengindentifikasi isolat NDV.
PENENTUAN GENOTIPE VIRUS
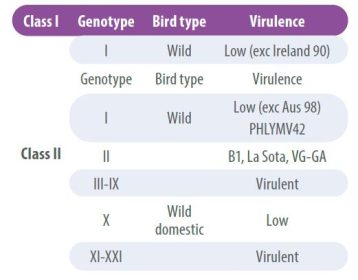
Virus NDV memiliki tingkat rekombinasi yang rendah, namun, seiring berjalannya waktu, perbedaan antigenik tertentu telah terdeteksi yang mengarah pada klasifikasi strain ke dalam garis keturunan atau genotipe. Oleh karena itu, strain dikelompokkan ke dalam dua kelas utama, virus kelas I dan virus kelas II;
- Virus kelas I memiliki patogenisitas rendah dan ditemukan pada burung liar.
- Di sisi lain, Kelas II diklasifikasikan ke dalam beberapa genotipe dan dapat bersifat nonpatogenik atau virulen (Tabel 2).
Karakterisasi genotipe NDV dilakukan dengan mengurutkan gen F secara lengkap, yang juga memungkinkan untuk menentukan virulensinya;
- Untuk mengidentifikasi genotipe baru, sebuah sistem yang didasarkan pada jarak evolusi rata-rata 10% antara kelompok genetik digunakan (Dimitrov KM et al 2016; Dimitrov KM et al, 2019).

Perkembangan sekuensing generasi berikutnya (NGS) telah muncul sebagai alat untuk penelitian dan karakterisasi genetik patogen.
- Namun, metode ini ideal untuk virus RNA yang bermutasi dengan cepat, di mana aspek evolusi juga perlu dianalisis (Butt et al., 2019).
Klasifikasi genotipe saat ini mencakup tiga genotipe baru (XIX, XX dan XXI), sehingga total ada 21 genotipe virus dalam virus kelas II (Dimitrov dkk. (2019).
RESISTENSI TERHADAP AGEN FISIK DAN KIMIA
Sebagai virus yang terbungkus, kelangsungan hidup avian Paramyxovirus mudah dihancurkan oleh agen fisik dan kimiawi seperti:
- Panas,
- Sinar ultraviolet,
- Proses oksidasi,
- pH dan
- Perlakuan dengan sebagian besar disinfektan.
Namun, publikasi tentang waktu bertahan hidup virus di luar inang menunjukkan perbedaan yang dikaitkan dengan suhu, kelembaban, dan lingkungan tempat virus dianalisis (Kinde et al., 2004).
PDF