


Bronquite Infecciosa das Galinhas: 90 Anos de Evolução


Trabalho agraciado com o III Prêmio DOWN de Veterinária.
Este artigo é dedicado in memorian aos Professores Osmane Hipólito e José Maria Lamas. A monografia “Bronquite Infecciosa das galinhas” agraciado do III Prêmio Down no Congresso Brasileiro de Avicultura, Belo Horizonte, Minas Gerais, 1979 reúne os trabalhos realizados que resultaram na liberação acertada da vacina H120 e H52 para o controle da BIG no Brasil. Justificam os autores:
.….decorridos 22 anos após isolamento e identificação do vírus no país (Hipólito, 1957) continuamos aguardando providências que permitam enfrentar o problema em bases técnicas….dar-nos-emos por satisfeitos se o trabalho ora apresentado puder ser de alguma valia para a valorosa classe dos avicultores a quem, na oportunidade, rendemos nossa humilde homenagem.
Vírus da bronquite infecciosa: o primeiro CoV, pandêmico desde 1940
Bronquite infecciosa das galinhas (BIG) foi originalmente descrita como sendo uma doença respiratória contagiosa de frangos de corte, notificada em 1931 nos EUA, causada por um vírus denominado de Massachusetts 82828 (ou M41 ou VR21), que se disseminou para a Europa e Oceania em 1940, por exportação de galinhas.
O vírus Massachusetts evolui para amostras com tropismo para o sistema gênito-urinário (e.g. amostra T, Cumming, 1940 Austrália e Holte-Gray ,1963, EUA) e mais de 50 variantes antigênicas que existem hoje no mundo.
As amostras US/Conn/Conn46/51 e H-52, 1955 Holanda (Bijlenga et al., 2010) foram as primeiras variantes introduzidas para controle da BIG. O vírus da BIG (IBV = Infectious Bronchitis Virus) foi o primeiro Coronavirus (CoV) descoberto no mundo, classificado somente em 1968.
Infectious Bronchitis Virus
O IBV é um patógeno economicamente importante para a indústria avícola porque infecta várias espécies de aves sinantrópicas e selvagens e só causa doença nas galinhas.

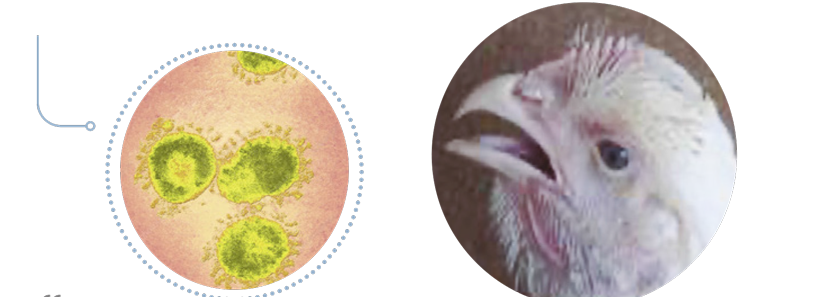
Bronquite Infecciosa das Galinhas
A inflamação nos condutos lacrimais e nasais causa espirros e coriza e quando associado com E.coli evolui para celulite perifacial. Ao início da BIG frangos apresentam estertor traqueal bolhoso e dificuldade inspiratória.


Bronquite Infecciosa das Galinhas
Frangos infectados com IBV apresentam:
- Queda de desempenho zootécnico;
- Espirros,
- Estertores traqueais e broncopneumonia,
- Aumento de mortalidade por diurese, uremia e nefrite nefrose,
- Comprometimento da eficiência alimentar por enterite associada com outros vírus (De La Torre et al., 2018) e multiplicação no íleo e favorecimento de aumento de OTUs de Enterobacteriaceae (Chu et al., 2020) e
- Condenação de carcaças por doença respiratória complicada (“cabeça inchada” e doença crônica respiratória ou DCR).
Galinhas de postura e reprodutoras adultas infectadas com IBV apresentam:
- Queda de produção e
- Produzem ovos com má qualidade de casca e albumina, e
Se infectadas no período de recria e/ou de crescimento:
- Têm produção limitada de ovos;
- Mesoperitonite por refluxo de ovos;
- Impactação de ovos no magno;
- Urolitíase, ou são falsas poedeiras
Os galos
Galos infectados com AvCoV apresentam testículos pequenos ou com atrofia cística dos túbulos seminíferos devido à replicação do vírus nas células epiteliais do epidídimo e obstrução dos dutos epididimários com células necrosadas e cálculos de cálcio ou oxalato.
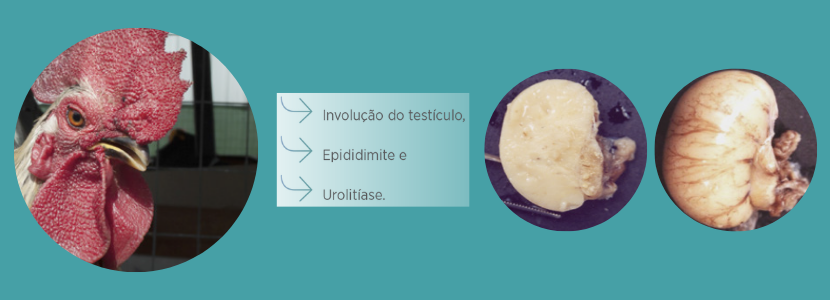
Bronquite Infecciosa das Galinhas
Galinhas infectadas com AvCoVs (Wille & Holmes, 2020) patogênicos produzem ovos com clara aquosa porque replicam nas células epiteliais do magno e com casca fina ou descolorida porque replicam nas células do istmo e útero. Galinhas infectadas com M. synoviae e AvCoV tem aumento da produção de ovos com defeito de calcificação no ápice. Galinhas com BIG produzem ovos sem casca ou pequenos, tipo cobra ou estrangulados, porque tem atrofia do istmo e útero ou aerossaculite abdominal ou mesoperitonite.

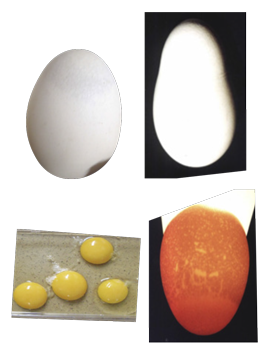
Bronquite Infecciosa das Galinhas
Taxonomia: lições e estratégias para biossegurança
Coronavírus (CoV) aviários são Nidovirales, família Coronaviridae, subfamília Orthocoronavirinae, conforme ICTV 2018, diferenciado conforme afinidade de hospedeiro, patogênese e composição genômica em 2 gêneros – Gammacoronavirus (GammaCoV) e Deltacoronavirus (DeltaCoV).
GammaCoVs
Os GammaCoVs são de três subgêneros:
1) Cegacovirus, representada pelo CoV SW1 da baleia Beluga e golfinho;
2) Brangacovirus com a espécie GoCoV (Goose Coronavirus) CB17 letal e específico de gansos e
3) Igacovirus, com três espécies – Avian CoV (AvCoV), Avian CoV 9203 e Duck CoV 2714 (Wille & Holmes, 2020).


Bronquite Infecciosa das Galinhas
AvCoVs são todos os vírus tipo IBV que Igacovirus infectam galinhas, perus, codornas, patos, gansos, pelicanos, papagaios, gaivotas, pombos, corvos e pardais, e inclui as amostras, PhCoV da doença respiratória em faisões e GfCoV, da enterite aguda e pancreatite em galinhas d´angola.
AvCoV 9203 é o vírus da enterite e síndrome da mortalidade aguda dos perus, que induz infecção assintomática em galinhas (Guy, 2010; Witt & Cook, 2020) e
DCoV 2714 patogênico para patos também circula entre os pombos, galinhas e gansos (Chen et al., 2013).
GammaCoVs mosaicos podem surgir por recombinação das três espécies. São candidatas a novas espécies de GammaCoV, CoVs exclusivos ou “dominantes” de patos e pombos com mutação do ORF poliproteína replicase (Zhuang et al., 2020).
DeltaCoVs
Os DeltaCoVs ocorrem em 15 ordens distintas de aves de pelo menos 30 famílias e 108 espécies de aves domésticas e selvagens, inclusive em codornas japonesas criadas comercialmente em consorciação com galinhas de postura (Torres et al., 2016).
DeltaCoVs são diferenciados em 3 subgêneros:
1) Andecovirus (Wigeon CoV HKU20),
2) Buldecovirus (Bulbul CoV HKU11, Common moorhen CoV HKU21, CoV HKU15, Munia CoV HKU 13, White-eye CoV HKU16) e
3) Herdecovirus (Night heron Cov HKU19).
Muitas amostras detectadas nas aves selvagens ainda não foram classificadas, e algumas delas, por exemplo, DeltaCoV da codorna e do papagaio Amazona virigligenalis, têm potencial para causar doenças nas aves comerciais.
Não existe risco de recombinação dos GammaCoVs com AlfaCoVs e BetaCoVs, porque as aves domésticas não são susceptíveis aos vírus dos mamíferos e vice-versa, contudo, GammaCoVs que infectam patos, codornas domésticas e aves selvagens, podem recombinar com DeltaCoVs.
DeltaCoVs foram detectados no gato leopardo e ferret. O DeltaCoV porcino (PDCoV), isolado em 2009 na Ásia, é um Buldecovirus recombinante CoV HKU15, com potencial para infectar células de humanos, galinhas e outras espécies de aves, porque usa o receptor aminopeptidase N (Wille & Holmes, 2020).
AvCoVs nunca serão extintos

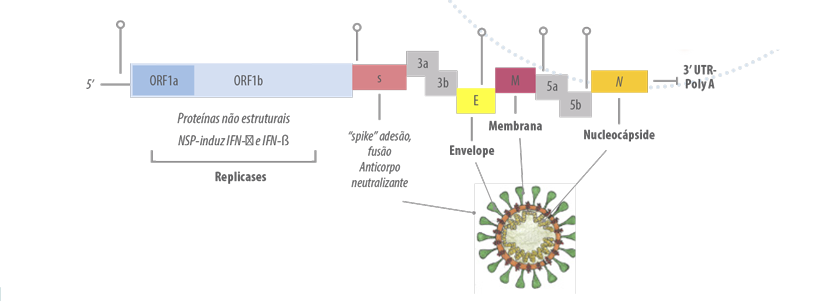
AvCoVs são virus +ssRNA com genoma de 27,5 a 28 kb e sequência gênica 5’ UTR-leaderORF1a/1b-S-3a 3b-E-M-5a 5b-N-3’UTR.
CoVs geram variantes genotípicas (e fenotípicas) por mutação, ou por recombinação intraespécie e interespécie (RNA quiméricos), que se disseminam e se fixam por seleção natural.
A mutação no ORF1ab que codifica as proteínas não estruturais, que resultam na formação do Nsp12, ou RNA polimerase, dependente de RNA viral (RdRp) e do Nsp 14 que tem a exonuclease da superfamília DEDD derivada do hospedeiro, induz erro de incorporação de nucleotídeos de até 10-4 a 10-5/ponto/ano nos AvCoVs, que é 10x mais elevada que a dos Retrovírus e 10.000 x mais elevada que a maioria dos vírus DNA (Legnardi et al., 2020).
AvCoVs mutantes são gerados por infecção natural e cultivo in vitro em células, ou em ovos embrionados de galinhas. Mutações não sinônimas só ocorrem nas aves vacinadas, porque a resposta imune é a principal força seletiva para evolução, por inversão de fase e deleção, ou inserção de nucleotídeos, ou aminoácidos nos determinantes antigênicos.
Vacinas
As vacinas vivas podem ter mais de uma subpopulaçao de vírus propensas a rápida mutação (Gallardo, Van Santen & Toro, 2010; Saraiva et al., 2018), reversão de virulência, ou seleção de variantes adaptadas (McKinlley, Hilt & Jackwood, 2008; Legnardi et al., 2020).
As vacinas vivas que induzem reação prolongada e elevada resposta de anticorpos têm elevada propensão para reversão de virulência, porque apresentam subpopulações de vírus que replicam em diferentes tecidos (Legnardi et al., 2020), ou podem induzir mutação de AvCoVs circulantes porque são muito imunogênicas, (McKinley et al., 2011).
AvCoVs efetuam recombinação mediada por RdRp quando se aplica múltiplas vacinas vivas ou quando o vírus vacinal circula por muito tempo na população, ou as aves tem imunodeficiência ou estão coinfectadas com Mycoplasma.
A vacina 793B efetua 3×10-3 substituições de aminoácidos por ponto/ano no gene S1 (Toro, Van Santen & Jackwood, 2012) e um AvCoV pode dar origem a variantes recombinantes inter-linhagem muito virulentos, tais como:
HN08, ou CK/CH/LSC/991 na China e Coreía (Valastro et al., 2016) e
Mais recentemente no Egito, ao IBV/CK/EG/Fadllh-10/2019 isolado de galinhas de postura hipersensibilizadas com Ma5 (Rohaim et al., 2019).
AvCovs são classificados em sorotipos por provas de soroneutralização com anticorpos monoclonais ou em genotipos neutralizantes por sequenciamento do gene que codifica o domínio S1 que tem três regiões hipervariáveis (HVRs):
- HVR1 (aa 60-88);
- HVR2 (aa 115-140) e
- HVR3 (aa 275-292)
Conforme Valastro et al. (2016), por análise de 1.286 sequências S1 completas catalogadas no GenBank no período de 1937 a 2010 (não incluídas 232 amostras recombinantes RdRp), existem 6 genótipos puros de AvCoVs:
- GI diferenciadas em 27 linhagens detectadas em diferentes países do mundo;
- GII na Holanda;
- GIII e GV na Austrália;
- GIV nos EUA; e
- GVI na China
A linhagem GI-1, que inclui AvCoV Beaudette e amostras derivadas das vacinas Van Roeckel M41 e H120 é cosmopolita, e a variante e GI-13 tipo vacina 793B detectada na Europa e na Ásia, não ocorre nos EUA, Oceania e muitos países da África e América Latina.
Segundo Valastro et al. (2016), os AvCoVs UFMG/G/1975, IBV/Bras/351/1984, UFMG/297 e 283, 1983 e UFMG/1141 e 12.2, 2009 catalogadas no GenBank por Abreu et al. (2006) são da linhagem S1 GI-11 exclusiva do Brasil.
Segundo Montassier et al. (2008), foram detectadas cinco linhagens distintas causando doença renal e respiratória no Brasil por sequenciamento de um fragmento RT-PCR/RFLP S1 de 1,72 kb de 12 AvCoVs isolados no período de 1988 a 2000, infelizmente ainda não diferenciadas segundo modelo de Valastro et al. (2016).


Bronquite Infecciosa das Galinhas
Amostras do tipo não Mass detectadas entre 2002 a 2006 (Villarreal et al., 2017), 2007 a 2008 (Villarreal et al., 2010), 2003 a 2009 (Felipe et al., 2010), 2010 a 2011 (Fraga et al., 2013) estudadas por análise parcial do gene S1, não podem ser designadas como novos genótipos ou linhagens S1 (Valastro et al., 2016).
Por reconstrução da árvore ML pelo modelo de Valastro et al, Fraga et al (2018) presumem que 24% das AvCoVs de material clínico do Brasil são GI-1 tipo Mass (24%), 74,5% são GI-11 (antes SA-I), 1% GI-13 tipo 4/91 e 0,5% GI-9 tipo Ark.
Patobiologia: epizootiologia e patogênese
AvCoVs são vírus sensíveis a temperatura elevada, radiação solar, pH extremo e desinfetantes, entretanto, sobrevivem nas fezes.
AvCoV se transmitem eficientemente por contato direto ave a ave, até a uma distancia de mais de 1,5 metros, e entre aviários, a uma distância de mais de 1.200 metros, favorecida pela direção dos ventos.
AvCoVs utilizam como porta de entrada a conjuntiva, seios nasais e o trato gastrintestinal. Efetuam replicação primária nas células epiteliais e glandulares do ducto lacrimal, seios nasais, traqueia, proventrículo e intestino delgado. A replicação secundária ocorre no rim, sistema reprodutor e outros órgãos tais como fígado, bolsa e baço.


Bronquite Infecciosa das Galinhas
Na fase aguda da BIG a mucosa da traquéia se apresenta edemaciada, úmida, discretamente amarelada e com brilho e a partir de 7 a 10 dias após-infecção, a mucosa fica delgada, seca e com aumento de área de epitélio com metaplasia cuboidal, e subsequentemente, presença de pontos brancos com halo avermelhado (ninho linfóide = BALT) se projetando para o lúmen, em quantidade e tamanho variável conforme patogenicidade do vírus, taxa de recontato e associação com outros patógenos respiratórios virais e micoplasma. Aves com traqueíte têm perda da defesa constitutiva muco-ciliar e sacos aéreos abdominais com exsudato inflamatório bolhoso ou complicado com E. coli, Ornithobacterium rhinotracheales, Pasteurella spp, etc.
O tropismo do AvCoV por células de diferentes tecidos do hospedeiro varia conforme quantidade de cópias de S1 que ligam no ácido siálico α-2,3, ou nos receptores do tipo lectina (Wickramasinghe et al., 2011) e composição de aminoácidos.
Os aminoácidos 19 a 272 do S1 ligam ácido siálico das células respiratórias, os aminoácidos 273 a 532 ligam substratos com açúcar (Neu 5 Aca2-3Gal) das células do oviduto, rim e intestino (Promkuntod et al., 2014) e os 110 a 112 ligam no rim (Bowman et al., 2020).
O S1 do M41 prefere ácido siálico com lactosamina tipo I [Gal(1-3)GlcNAc] que o do tipo II [Gal(1-4)GlcNac], que predomina na traqueia, rim, sacos aéreos, seios nasais, íleo, intestino grosso, bolsa de Fabrícius e capilar do pulmão das galinhas (Wickramasinghe et al., 2015).

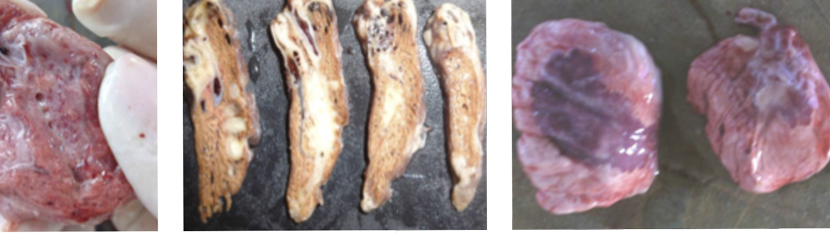
Bronquite Infecciosa das Galinhas
AvCovs patogênicos causam edema nos brônquios primários e secundários e formação de um tampão de células mortas e inflamatórias que causam obstrução do lúmen dos mesobrônquios secundários e terciários. A obstrução dos brônquios e mesobrônquios médio-ventrais resulta em enfisema pulmonar e broncopneumonia congestiva focal visível macroscópicamente na região costal do pulmão (paleopulmão)
A capacidade de adesão e fusão do AvCoV na célula epitelial também depende da presença de determinados aminoácidos na posição 38 e 43 proteína S1: Ma5 (Brandão et al., 2017) e M-41, QX e Conn 46, têm tropismo para traqueia porque têm asparagina na posição 38. A presença de histidina na posição 43 da M-41 aumenta a afinidade, sendo a substituição destes aminoácidos responsável pelo baixo tropismo da H120, ou abolição da adesão na amostra Beaudette (Promkuntod et al., 2014).
AvCoV nefropatogênica B1648 (Reddy et al., 2016), M41, Cal 99, Conn 46 e Iowa 97 replicam em monócitos/macrófagos circulantes (Zhang & Whittaker., 2016) e impedem a produção de óxido nítrico (Amarasinghe et al., 2017) para se disseminar para o fígado, rim, baço, pulmão e outros tecidos.
Galinhas sem imunidade permanecem portadoras por longo tempo (2 a 7 meses) e o vírus é excretado nas excreções oculares e nasais, fezes, urina, sêmen e ovo. AvCoVs podem ser detectados na membrana vitelínica do ovo por 1 a 7 semanas após desafio, no sêmen por até 2 semanas (Jackwood & de Witt, 2019) e no fluido alantoide de ovos embrionados (Pereira et al., 2016).
AvCoVs nefropatogênicos causam inicialmente aumento de volume do lóbulo renal devido à degeneração das células epiteliais, distensão e retenção de uratos no lúmen dos túbulos contornados e infiltração focal de células mononucleares resultando em eliminação profusa de fluidos e íons na urina (diurese), miopatia peitoral e morte por desequilíbrio ácido básico.

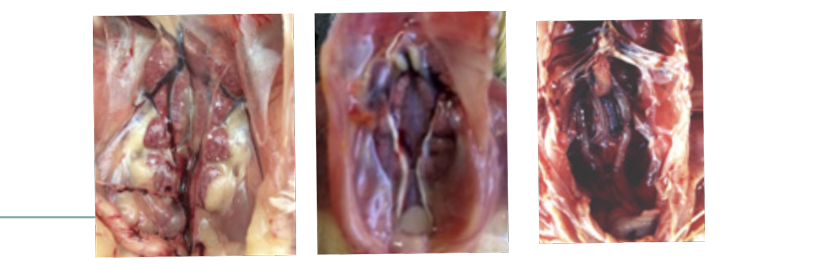
Bronquite Infecciosa das Galinhas
Pintinhos com nefrite nefrose apresentam desidratação, ureteres com retenção de uratos e enterite (íleo > jejuno). AvCoVs nefropatogênicos são excretados na urina e detectados em suabes de cloaca e/ou de reto.
Aves que sobrevivem apresentam atrofia dos lóbulos renais e galinhas de postura e reprodutoras infectadas durante a recria apresentam urolitiase tubular e ureteral ao serem alimentadas com ração pré-postura com mais cálcio.
A imunidade induzida pelas vacinas vivas e inativadas AvCoVs não impede a reinfecção, mas reduz significativamente o tempo e taxa de replicação do vírus desafiante homólogo.
Diagnóstico e controle: ponto crítico
O diagnóstico sorológico da BIG é dificultado quando as aves:
- Recebem múltiplas vacinas;
- Apresentam coinfecção com outros patógenos (principalmente Mycoplasma gallisepticum e M. synoviae);
- Apresentam imunodeficiência e
- São utilizadas vacinas que favorecem circulação de vírus.
Por isto, para confirmação de caso suspeito deve-se efetuar exame virológico (isolamento e PCR) e histológico de múltiplos órgãos coletados durante fase aguda da doença.
Os métodos de diagnóstico baseados em RT-PCR genérico (e.g. 3’ ou 5’ UTR ou S1) feito pelo método Sanger, que detecta somente uma amostra (geralmente a de título elevado), e RT-PCR(qRTPCR) específicos (Legnardi et al., 2020) para uma a duas linhagens, não permitem identificar a diversidade de AvCoVs que estão circulando entre a aves comerciais e sinantrópicas.
Por exemplo, o RT-PCR(qRT-PCR) específico para fragmento S1 325 a 443 H-120 e BR-I não permitiu identificação de 55/141 amostras clínicas positivas no RT-PCR(qRTPCR) para 5’UTR AvCoV (Fraga et al., 2016).
Para sabermos como AvCoVs evoluem é preciso conhecer a dinâmica do genoma completo dos vírus vacinais e de campo nos diferentes compartimentos funcionais do hospedeiro e integrar os resultados com o padrão de resposta de anticorpos, alterações microscópicas e manifestações clínicas em diferentes nichos ecológicos.

Prevenção
A prevenção da BIG deve ser baseada em:
Escolha de vacinas com espectro limitado de disseminação e persistência endógena e transmissibilidade intra-espécie e interespécie de aves;
Condições de conforto térmico;
Controle de patógenos respiratórios como micoplasma ou imunossupressores;
Condição de biosegurança, por exemplo, esxistencia de aves que sirvam de reservatrios; e
Vigilância por estudo longitudinal por RT PCR e qRT-PCR genérica, ou específica e sorologia.
O Brasil é um dos poucos países que tem poucas linhagens definidas por análise da sequência de nucleotídeos no gene S1, além da GI-1 tipo Mass e GI-11.
Talvez porque desde 1977 (Portaria Divisão de Defesa Sanitária Animal nº 003, 13/05/77) a vacina H-120 que atende todos os critérios para imunização ativa de pintinhos (Bijlenga et al., 2010), é a vacina mais usada no país.
Ainda não sabemos qual é a relação do grau de adaptação do vírus em um indivíduo, com o restante de indivíduos de uma determinada população de galinhas e porque algumas variantes ficam restritas à uma área geográfica e ressurgem periodicamente, enquanto outras são capazes de persistir e disseminar para o mundo inteiro.
O genoma completo da H120 é similar da H52, ARK DPI, Beaudette, M41 e ZJ 971, exceto que, H120 e H52 tem 3’UTR mais intacta e longa (Zhang et al., 2010) e não tem asparagina e histidina respectivamente nas posições 38 e 43 da molécula S1 (Pronkuntod et al., 2014).
A vacina H52, que tem uma grande quantidade de subpopulação de vírus instável (Bijlenga et al., 2010) foi retirada do mercado brasileiro em meados de 1990 por ser muito virulenta e subsequentemente, H-72 e H-90. A partir de 2000 foram introduzidas Ma5 e B48.
A introdução da vacina IBRAS derivada da amostra BR-1 GI-11, que existe no país desde antes de 1975 (UFMG/1975) e circula no país desde 1950 (Fraga et al., 2018), deve mudar a dinâmica do AvCoV.

A BR-I tem tropismo para trato digestivo, reprodutor e respiratório das galinhas (Balestrini et al, 2014), causa discreta a moderada degeneração das células epiteliais da traquéia da galinha SPF (Chacon et al., 2014; Santos et al., 2017) e é mais severa no sistema respiratório de frangos (Chacon et al., 2014).
A cinética e intensidade das lesões microscópicas induzidas pela BR-I são equivalentes à da H-120 (Katayama Ito, comunicação pessoal), que não causa destruição do epitélio traqueal entre 1 a 7 dias após infecção experimental por aerolização em pintinhos SPF com o M41 e 2814/89 isolada de rim de frango com nefrite nefrose.
A amostra H-120 causa moderada traqueite linfoproliferativa entre 3 a 7 dias após infecção à semelhança das amostras 2370/89 e 3033/77 isolada de rim, respectivamente de galinhas d’angola e frangos com nefrite nefrose (Miyaji, 1996).
Hoje sabemos que CoVs são vírus que matam as células por esgotamento metabólico ou que favorecem apoptose porque induzem uma resposta imune vigorosa mediada por células (TCD8 e TCD4Th1).