



A bronquite infecciosa das galinhas (BIG) é doença aguda altamente contagiosa de frangos e galinhas (Gallus gallus domesticus).
A BIG é considerada atualmente uma das doenças que mais causa prejuízos à avicultura industrial de galinhas e frangos no Brasil e no mundo.
Há grande impacto econômico, por causar aumento da mortalidade, perdas em desempenho produtivo, queda quantitativa e qualitativa na produção de ovos, infertilidade, facilitação de infecções oportunistas, que exigem o uso de antibióticos, e aumento na condenação de carcaças de frangos de corte no abatedouro [2,5]. É importante destacar que, em codornas domésticas (Coturnix japonica e Coturnix coturnix), a bronquite é causada por um adenovírus (Aviadenovirus), sem qualquer relação com a etiologia de BIG.
Etiologia
A BIG é causada pelo vírus da bronquite infecciosa (IBV – infectious bronchitis virus), classificado no gênero Gammacoronavirus, família Coronaviridae, ordem Nidovirales, com genoma de RNA fita simples e polaridade positiva, e com envelope lipoproteico [2, 5, 9]. Os coronavírus descritos em faisão, galinha-d’angola, marreco e peru têm semelhança genética com IBV [3, 4, 11, 12]. Uma importante
característica de IBV é a grande diversidade genética e antigênica.
Uma população de IBV nunca é geneticamente idêntica, e a heterogeneidade (quasispecies) é a principal estratégia evolutiva de evasão à resposta imune e dificulta a eficiência vacinal. Outra estratégia evolutiva de IBV é a possibilidade de rearranjo genético entre diferentes
IBVs [5, 9], podendo dar origem a progênies virais de risco para plantéis com proteção prévia heteróloga. O IBV foi o primeiro coronavírus descrito em animais (1937), nos Estados Unidos [5].
As estirpes de IBV podem ser classificadas em sorotipos heterogêneos com baixa proteção cruzada [2, 5], característica que torna difícil a implantação de programas eficientes e duradouros de vacinação.
No envelope de dupla camada lipídica, as grandes projeções superficiais (S) são glicoproteínas essenciais para infecção, com papel na
adsorção e penetração na célula. A região mais externa de S, denominada S1, liga-se aos receptores celulares glicoproteicos de tipo ácido siálico. Variações na composição de aminoácidos em regiões hipervariáveis de S1 determinam diversidade antigênica e de sorotipos [2, 5]. O
RNA é copiado na célula infectada em seis segmentos de RNA subgenômicos (mensageiros), os quais, em coinfecção entre IBVs diferentes e na montagem de novos vírions, podem gerar recombinantes, com potencial de evasão à imunidade preestabelecida, representando o principal fator na emergência de novo surto de BIG em plantéis vacinados.
No Brasil, em Minas Gerais, há estudos que demonstram a presença de IBVs com amplas variações genéticas [1] e sorológicas [7, 14, 15] na avicultura industrial, além de relatos também em outros estados, como SP [16], SC, PR e RS, sendo atualmente descrito um grupo brasileiro (BRI).
Epidemiologia
A bronquite infecciosa das galinhas é causada por IBV e ocorre naturalmente em Gallus gallus domesticus. Em faisões e perus ocorrem coronavírus assemelhados e não IBV. Coronavírus do peru (TCoV) e coronavírus do faisão (PhCoV) são espécies distintas de IBV dentro de Gammacoronavirus, com apenas 34%, em média, de identidade no gene que codifica a glicoproteína S1. Estirpes de IBV mantêm entre si 75 a 85% de identidade em S1 [3, 11]. Entretanto, isolados de IBV semelhantes à estirpe vacinal H120 (>99% de identidade em S1) foram obtidos de pavão e marreco assintomáticos [12].
Um episódio de doença respiratória foi descrito em galinha-d’angola, com isolamento de coronavírus relacionado antigenicamente (neutralização cruzada) com IBV. Anticorpos de galinhas matrizes vacinadas contra IBV foram neutralizantes contra o isolado [10]. Uma diversidade de Gammacoronavirus foi detectada em espécies de aves domésticas e silvestres, das ordens Ciconiiformes, Pelecaniformes e
Anseriformes [3].
Os estudos mais recentes indicam que a epidemiologia de coronavírus é complexa em classe Aves. Deve-se manter a mente aberta para a possibilidade de que a existência de múltiplos hospedeiros para infecção por IBV pode, eventualmente, resultar em doença em novas espécies [3].
Transmissão
A transmissão ocorre principalmente por aerossóis respiratórios, com vírus replicado no epitélio respiratório e eliminado em microgotas de muco respiratório durante a expiração [2, 5].
Nas regiões de concentração da avicultura (de Gallus gallus domesticus), há problemas continuados com BIG, pela proximidade e fácil difusão aérea de IBV em aerossóis respiratórios. Em granjas, ou regiões, com ocupação contínua de frangos de corte ou galinhas (sem vazio sanitário ou com falhas de biosseguridade), a imunidade induzida por vacinação pode eventualmente ser superada por variantes virais.
Nessas condições, de proximidade e falha de biosseguridade, com presença contínua de aves, a transmissão é fácil, e a erradicação difícil. Como agravante, as estirpes que circulam na região podem ser derivadas de estirpes vacinais, por mutação, ou recombinação com IBVs de campo.
Entretanto, é possível o controle de BIG em granjas ou núcleos sem vacinação contra BIG, que mantenham programa de biosseguridade, com manejo exclusivo, distanciamento, idade única e vazio sanitário entre lotes. A doença pode atingir os sistemas respiratório, reprodutor, gastrointestinal e/ou excretor, em galinhas ou frangos, sendo recorrente tipicamente em granja ou região densamente povoada por múltiplas idades. O IBV infecta os epitélios respiratório, gastrointestinal, renale do oviduto, em intensidade e impacto variável conforme a estirpe [2,5].
Há predominância de estirpes mais adaptadas à infecção respiratória, embora com reflexos nos demais sistemas citados, excretor (rins e ureteres) e reprodutivo (regiões do oviduto, magno e útero).
Patogenia
Para a infecção das células-alvo no epitélio ciliado respiratório, gastrointestinal, tubular renal, no magno e no útero, as glicoproteínas (S1) no envelope viral de IBV adsorvem aos receptores celulares que contêm ácido siálico, e essa atividade é dependente da integridade do envelope [2, 5].
A infecção respiratória é mais grave em aves mais jovens, com possibilidade de obstrução traqueal parcial (Fig. 1) ou total por muco, seguida de asfixia e morte. A infecção renal ocorre tipicamente após a infecção respiratória, exceto para estirpes de infecção renal exclusiva. Alguns IBVs produzem lesões graves nos rins (nefrite-nefrose), com morte por insuficiência renal.
A infecção do oviduto nas fêmeas maduras e em produção resulta em, no magno, falha na síntese das proteínas do albúmen e, no útero, falha na produção da casca do ovo. A lesão no magno se caracteriza por produção de ovos com clara aquosa, e a lesão no útero por ovos de casca mole, frágil, mais fina e despigmentada, podendo também ocorrer ovos com deformidades na casca.
A BIG apresenta gravidade variável com a estirpe de IBV envolvida, idade da ave, genética, sexo, qualidade do manejo, do ambiente, doenças concomitantes e status imune. Os problemas mais graves ocorrem em granjas que mantêm populações de galinhas/frangos de idades
variadas e próximas. O desafio de pintinhos nas proximidades de aves mais velhas, especialmente para frangos de corte e poedeiras em recria, pode resultar em quadros mais graves.
A BIG é tipicamente agravada por Escherichia coli, bactéria inalada em suspensão no ar, em galpões com baixa renovação de ar e alta concentração de poeira.
Quadros respiratórios graves ocorrem, com maior frequência, em criações com baixa qualidade sanitária e de manejo, em galpões com baixa renovação do ar, tipicamente para aves que estão no chão, como frangos e corte e poedeiras em cria [2, 5].
Sinais clínicos
A doença predominante é respiratória (Fig. 1), com estertores, tosse, dispneia, insuficiência respiratória, asfixia e morte dos mais jovens.


Figura 1: Galinha com insuficiência respiratória e respiração com o bico aberto. Notar a presença de conjuntivite mucoide. Assista ao vídeo no YouTube
Os estertores respiratórios resultam da ciliostase e acumulação de muco no lúmen traqueal (veja a atividade ciliar normal no vídeo abaixo).

Nas aves jovens, a forma respiratória pode evoluir para a forma renal, com a infecção que destrói a estrutura e a função renal, levando à mortalidade por insuficiência renal.
Na traqueia, pode haver muco (Fig. 2), congestão e hemorragia, e nos rins acúmulo de uratos nos mesonefrose ureteres, com aumento de volume e marmorização (Fig. 4).

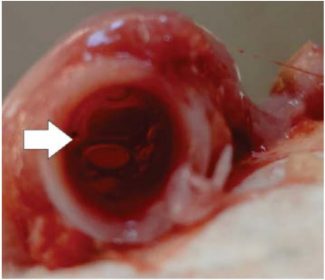
Figura 2. Imagem de traqueia de ave (Gallus gallus domesticus) jovem com muco espumoso em obstrução parcial do lúmen (seta).

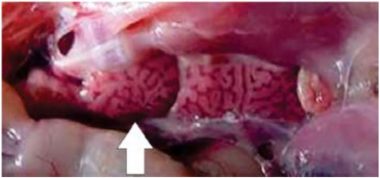
Figura 4. Processo de nefrite-nefrose renal (seta)
Muitos IBVs podem infectar o oviduto de galinhas em crescimento ou em postura. As lesões no magno e no útero podem ser irreversíveis e resultar na formação de ovos com anomalias na clara e na casca, com cascas frágeis, trincadas, irregulares, moles ou ovos sem casca (Fig. 3).
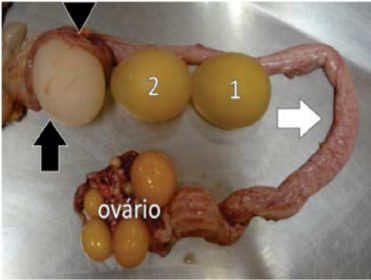
Figura 3. As regiões do oviduto atingidas, magno (seta branca) e útero (seta preta), resultam, respectivamente, em clara aquosa e ovos de casca frágil, mole ou sem casca. Notar, no útero aberto, um ovo em formação de casca (ponta de seta). Dois ovos sem casca, com uma (1) ou duas (2)
membranas de casca, estão enfileirados sobre
a região onde são produzidas essas membranas (istmo).
Alguns IBVs infectam os intestinos e resultam em diarreia, desidratação, deficiente digestão, absorção de nutrientes e nanismo infeccioso, sendo normalmente esses quadros associados a outros patógenos, como rotavírus, reovírus, calicivírus, parvovírus, entre outros, sugerindo ser um quadro multicausal [2, 4, 5].
Diagnóstico
A suspeita é especialmente significativa se há histórico de ocorrência prévia de BIG e não houve vazio sanitário ou não se empregaram estratégias adequadas de biosseguridade. Entre as viroses respiratórias, a BIG é a mais comum em regiões com alta densidade avícola,
proximidade de granjas e multiplicidade de idades. Há doenças de manifestação clínico-patológica semelhantes à BIG, como as micoplasmoses (Mycoplasma gallisepticum, M. synoviae) e as colibaciloses (Escherichia coli), que podem ocorrer como coinfecções com
IBV, pneumovirose (metapneumovírus aviário), laringotraqueíte infecciosa (Iltovirus) e doença de Newcastle (Avulavirus) por estirpes mesogênicas ou lentogênicas.
O diagnóstico definitivo de BIG depende de confirmação laboratorial. Anticorpos específicos para IBV podem ser determinados por sorologia em ELISA [13] ou inibição da hemaglutinação (HI) [2,4,7]. Em aves vacinadas, podem-se observar títulos ascendentes
em duas coletas de soros, com intervalo de 15 a 21 dias, e, em aves não vacinadas, os títulos de anticorpos mínimossignificativos (acima de 8 em HI).
Para o diagnóstico definitivo, o vírus deve ser isolado e caracterizado em laboratório [2, 5].
O isolamento pode ser feito por cinco passagens consecutivas em ovos embrionados de nove a 11 dias de incubação, obtidos de galinhas SPF, incubados por até sete dias após a inoculação. Cultivos de anéis de traqueia podem ser usados para o isolamento e estudo de IBV.
Os tecidos de aves doentes ou anexos ou tecidos dos embriões podem ser examinados por métodos imunoenzimáticos ou por RTPCR
(transcrição reversa e reação em cadeia pela polimerase), para a detecção do IBV ou de seu genoma, respectivamente, geralmente após 48h de infecção em até cinco passagens.
A RT-PCR, seguida de análise dos produtos por restrição enzimática ou sequenciamento, permite a caracterização do isolado. A RT-PCR e a análise do produto amplificado podem também ser feitas a partir dos tecidos das aves doentes.
Prevenção e controle
A infecção natural depende da integridade do envelope de IBV. Entretanto, o envelope viral pode ser destruído por limpeza com detergentes e desinfetantes, assim como fatores naturais de inativação, como o calor e a radiação solar. Para a prevenção de BIG, pode-se necessitar do despovoamento (vazio sanitário) do núcleo, granja e/ou região, para a adequada desinfecção.
A uniformização das idades das aves no núcleo, granja ou região, pode permitir a adoção de vacinação geral na granja, infecção vacinal homogênea, em data única, potencialmente a melhor estratégia para proporcionar uniformidade de respostas e menor escape de vírus mutantes.
A resposta imune subprotetora é considerada indutora de pressão de seleção de vírus mutantes resistentes à imunidade prévia. A desuniformidade de status imune entre indivíduos em um plantel ou entre os plantéis pode resultar na emergência de IBV variantes, e perpetuam-se os episódios com vírus variantes.
A adoção de data única de vacinação pode ser importante estratégia, com o objetivo de reduzir o escape de vírus para plantéis sensíveis e uniformização de resposta imune. As tentativas de controle por vacinação em granjas, núcleos ou regiões que mantenham idades múltiplas podem ter efeito benéfico por alguns ciclos de produção, com diminuição progressiva da eficiência devido à variação viral.
No Brasil, para vacinas vivas, estão permitidas estirpes do sorotipo Massachusetts (H120, MA5, etc.), atenuadas por passagens consecutivas
em ovos embrionados de galinhas SPF. As vacinas vivas podem ser aplicadas em pintinhos após a eclosão, ainda no incubatório, por nebulização, ou na chegada à granja, por nebulização ou gota ocular. A via água de bebida (vacinas vivas) tem resultado em resposta imune desuniforme.
Para a proteção durante a produção de ovos, poedeiras e reprodutoras são geralmente revacinadas com vacina inativada aplicada via intramuscular no peito, que induz proteção sistêmica, especialmente importante para o oviduto. As vacinas inativadas, geralmente em emulsão oleosa para lenta absorção, são comumente combinadas para a proteção contra outras etiologias.
As complicações bacterianas são comuns em BIG e aumentam a morbidade e a mortalidade para aves mantidas em piso, frangos de corte e frangas de reposição, mantidas em ambientes com baixa renovação de ar e com poeira em suspensão, que facilita a infecção respiratória por
Escherichia coli. As infecções bacterianas secundárias comumente exigem terapia antibacteriana, escolhida com base em teste de sensibilidade.
Referências
1. Abreu, Josiane T.; Resende, J. S.; Flatschart, R. B.; Flatschart, Á. V. Folgueras; Martins, N. R. S.; Resende, M. Molecular Analysis Of Brazilian Infectious Bronchitis Field Isolates By Reverse Transcription Polymerase Chain Reaction, Restriction Fragment Length Polymorphism, And Partial Sequencing Of The N Gene. Avian Diseases V. 50, N.3, P. 494-501, 2006.
2. Avian Infectious Bronchitis, C H A P T E R 2 . 3 . 2 . OIE Terrestrial Manual 2013, Version adopted by the World Assembly of Delegates of the OIE in May 2013.
3. Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathology, 34, 439-448, 2005.
4. Chu, Daniel K.W.; Leung, C.Y.H.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. 2011. Avian coronavirus in wild aquatic birds. J Virol 85:12815–12820.
5. Cavanagh,, David; Naqi,, A.S. (2003) Infectious Bronchitis. In: Saif YM, Barnes HJ, Glisson JR, Fadly AM, Dougald MC, Swayne DE (eds) Disease of Poultry, 11th ed. Iowa State University Press, Ames, pp 101–119.
6. de Wit, J. J. (Sjaak), Cook, J. K. A. & van der Heijden, H. M. J. F.. Infectious bronchitis virus variants: a review of the history, current situation and control measures, Avian Pathology, 40:3, 223-235, 2011.
7. Epiphanio, Ericka.O.B.; Martins, N.R.S.; Resende, J.S.; Pinto, R.G.; Jorge, M.A.; Souza, M.B.; Caccioppoli, J.; Cardozo, R.M. Resultados preliminares da utilização de cultivos de anéis de traqueia para o estudo de estirpes brasileiras do vírus da bronquite infecciosa das galinhas. Arquivo Brasileiro de Medicina Veterinária e Zootecnia , Belo Horizonte, v. 54, n.2, p. 212-216, 2002.
8. Hewson, Kylie A.; Noormohammadi, A. H.; Devlin, J. M.; Browning, G. F.; Schultz, B. K. & Ignjatovic, J. Evaluation of a novel strain of infectious bronchitis virus emerged as a result of spike gene recombination between two highly diverged parent strains. Avian Pathology 43, p. 249-257, 2014.
9. ICTV. International Committee on Taxonomy of Viruses. 2002. http://ictvdb.bio-mirror.cn/Ictv/fs_coron.htm
10. Ito, N.M.K., Miyaji, C.I. & Capellaro, C.E.M.P.D.M. (1991). Studies on broiler’s IBV and IB-like virus from guinea fowl. In E.F. Kaleta & U. HeffelsRedmann (Eds.), II International Symposium on Infectious Bronchitis (pp. 302/307). Giessen: Justus Leibig University.
11. Lin, T.L., Loa, C.C. & Wu, C.C.. Complete sequences of 3’ end coding region for structural protein genes of turkey coronavirus. Virus Research, 106 , 61 /70, 2004.
12. Liu, S., Chen, J., Chen, J., Kong, X., Shao, Y., Han, Z., Feng, L., Cai, X., Gu, S. & Liu, M. (2005). Isolation of avian infectious bronchitis coronavirus from domestic peafowl (Pavo cristatus) and teal (Anas). Journal of General Virology, 86 , 719 /725.
13. Martins, N. R. S.; Mockett, A. P. A. ; Barrett, A. D. T. ; Cook, Jane K. A.. IgM Responses in Chicken Serum to Live and Inactivated Infectious Bronchitis Virus Vaccines. Avian Diseases, v. 35, p. 470, 1991.
14. Mendonça, Juliana F. P.; Martins, N. R. S.; Carvalho, L. B. de; Sá, M. E. P. de; Melo, C. B. de. Bronquite infecciosa das galinhas: conhecimentos atuais, cepas e vacinas no Brasil. Ciência Rural (UFSM. Impresso), p. 1-0, 2009.
15. Souza, M.B.; Martins, N.R.S.; Resende, J.S. Afinidades Antigênicas De Amostras De Campo Do Vírus Da Bronquite Infecciosa Das Galinhas Com A Amostra Massachusetts M41. Arquivo Brasileiro De Medicina Veterinária E Zootecnia, V. 53, N. 2, P. 1-7, 2001.
16. Villarreal, L.Y., Sandri, T.L., Souza, S.P., Richtzenhain, L.J., de Wit, J.J., Brandao, P.E. Molecular epidemiology of avian infectious bronchitis in Brazil from 2007 to 2008 in breeders, broilers, and layers. Avian Diseases, 54, 894-898, 2010.
Material retirado de “Cadernos Técnicos de Veterinária e Zootecnia, nº 76 – março de 2015”