


Como prevenir a imunossupressão e solucionar a questão do gap imunológico observados nos programas de vacinação contra Gumboro? Saiba mais aqui!
Uma doença aparentemente nova em galinhas tem sido frequentemente observada. Essa doença parece ser infecciosa e contagiosa, caracterizada por penas arrepiadas, diarreia e prostração. Os primeiros surtos ocorreram próximo à área de Gumboro, no sul de Delaware, Estados Unidos.
(Cosgrove, 1962)



Hoje sabemos que essa doença é causada por um vírus altamente contagioso, que afeta principalmente o sistema imune do seu hospedeiro.


No Brasil, surtos graves de Gumboro foram relatados desde julho de 1997 (Di Fabio et al., 1999). A doença com manifestações clínicas esteve muito presente na produção de frangos de corte e poedeiras até o surgimento de métodos imunoprofiláticos mais eficientes, como o uso de vacinas não convencionais, seja ela em sua forma vetorizada ou de complexo imune.


![]()
![]()
![]()
![]()
![]()

Considerando os efeitos redutores na população de células B que o vírus de Gumboro e que algumas vacinas convencionais exercem sobre o hospedeiro, sabe-se que as respostas imunes contra vírus e bactérias são negativamente influenciadas tanto em situações de desafio como também como efeito colateral com o uso de vacinas intermediárias e intermediárias plus4,5.
![]()


A depleção de linfócitos B na bursa de Fabricius após infecção é devida tanto à necrose quanto à apoptose (Hirai, 1979; 1981; Rodenberg et al., 1994).
QUAL O IMPACTO DA IMUNOSSUPRESSÃO CAUSADA PELO VÍRUS DE GUMBORO?

De modo geral, esse órgão fornece um microambiente para o desenvolvimento de linfócitos B, reconhecidos como as células responsáveis pela produção de anticorpos.


A imunossupressão causada pela bursectomia química, infecções pelo vírus da anemia infecciosa das galinhas e pelo vírus de Gumboro sobre a resposta imune contra a Salmonella Enteritidis e a Salmonella Typhimurium geram efeitos deletérios ao ambiente da bursa com efeitos persistentes e negativos tanto à resposta humoral quanto celular (Phillips e Opitz, 1995; Sheela, 2003; Wyeth,1975).
A respeito do uso de vacinas vivas convencionais de Gumboro tipo intermediária plus, tem-se mostrado que a vacina teve um papel importante ao desencadear um efeito deletério na resposta humoral contra a Salmonella Enteritidis, sendo que a resposta em relação à títulos de anticorpos séricos e de mucosa direcionados contra a Salmonella Enteritidis foi menor em grupos vacinados e desafiados quando comparado apenas ao grupo desafiado (Arafat et al., 2017).
![]()
Esse estudo aponta que, da mesma forma que ocorre com os vírus de campo, os vírus vacinais do tipo intermediário plus comprometem também a resposta celular e aumentam a fragilidade das respostas imunes para a defesa contra outros agentes infecciosos, como as salmonelas.

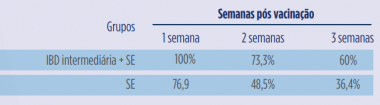
Tabela 1. Recuperação bacteriana de Salmonella Enteritidis em suabe de cloaca de aves vacinadas com uma vacina de gumboro tipo intermediária e desafiadas com Salmonella Enteritidis (adaptado de Arafat et al., 2017).
PREVENÇÃO E CONTROLE


No entanto, mesmo com boas condições de biosseguridade e aplicação do sistema “all-in all-out”, onde há a redução da carga viral e pressão de infecção na granja, o vírus ainda pode persistir no ambiente.


O primeiro conceito “pós vacinas convencionais” foi demonstrado com as vacinas de complexo imune.
Estas vacinas consistem em uma mistura de soro contendo anticorpos neutralizantes e específicos para o vírus de Gumboro, combinados com um vírus vivo, em geral estirpes intermediárias plus.
Essas vacinas são uma forma associada “antígenoanticorpo”, suficientemente equilibrada de modo a não neutralizar o vírus vacinal, mas capaz de retardar os efeitos patogênicos do vírus vacinal quando comparado à sua aplicação isolada.
No entanto, da mesma forma que as vacinas convencionais induzem lesões de Bursa, depleção linfoide e consequente imunossupressão, vacinas de complexo imune geram escores variados de lesão após a vacinação (Cazaban et al., 2018; Sedeik et al., 2019), com efeitos imunossupressores semelhantes às vacinas convencionais, sendo seu efeito mais tardio e dependente da queda dos títulos dos anticorpos maternos recebidos pela progênie.
Vacinas de complexo imune são administradas no incubatório, no entanto as estirpes presentes nestas vacinas não replicam de forma eficiente até que os anticorpos maternos atinjam um certo nível (Jeurissen et al., 1998), que fica abaixo do nível de proteção contra os vírus de campo.


Há, portanto, um período pós vacinal em que as aves são susceptíveis à infecção por vírus de campo. Este período é denominado “gap imunológico” (Fig. 1a).

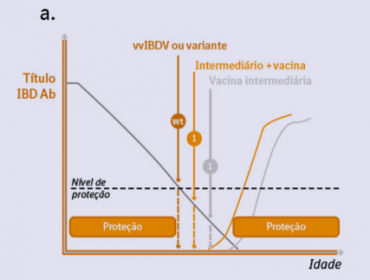
Figura 1a. Gap imunológico pode ser observado em vacinas vivas convencionais e de complexo imune.

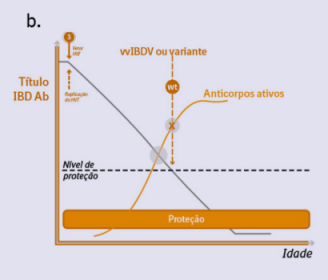
Figura 1b. Ausência de gap imunológico em aves vacinadas com vacina vetorizada HVT+IBD.
Outro modelo de imunoprofilaxia para a doença de Gumboro é representado pelas vacinas vetorizadas recombinantes, baseado na inserção do gene que sintetiza a proteína VP2 do vírus de Gumboro dentro do genoma do vírus vacinal de Marek do sorotipo 3 (HVT).
![]()

Esta proteína, em combinação com as proteínas do vírus de Marek, induzem uma resposta imune que protege a ave vacinada contra o vírus de Gumboro e o vírus de Marek.
As células do pintinho que contém a vacina HVT+IBD em seu interior passam a liberar uma nova progênie do vírus vacinal, que então entrará em contato com novas células alvo, repetindo esse ciclo sucessivamente e induzindo uma resposta imune sistêmica (Fig. 2), replicando-se em diversos órgãos da ave ao invés de apenas os linfócitos B, como as vacinas vivas convencionais e de complexo imune (Prandini et al., 2008; Bublot et al., 2007)

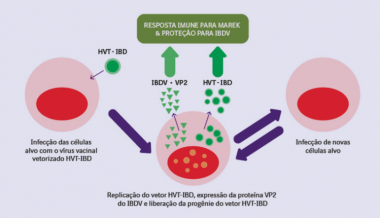
Figura 2. Interação do vírus da vacina HVT+IBD com o sistema imune das aves.
A vacina HVT+IBD possui o vetor do vírus de Marek com replicação rápida, sendo que sua replicação inicial se dá logo após a aplicação no incubatório, mesmo em aves com altos níveis de anticorpos maternais.
A replicação precoce do HVT e a expressão do gene VP2 de Gumboro permitem a indução precoce de anticorpos ativos contra o vírus de Gumboro, que compensarão progressivamente a queda do nível de anticorpos maternos, mantendo os níveis de anticorpos acima do limiar de proteção (Fig. 1b) (Bublot, 2015).
A tecnologia de vacinas vetorizadas, portanto, é a única que pode solucionar as questões de prevenção da imunossupressão e gap imunológico observadas nas vacinas convencionais e de complexo imune.

![]()


![]()
Diante da disponibilidade de diferentes tecnologias no mercado, ressalta-se a importância do médico veterinário em conhecer a situação epidemiológica da doença de Gumboro de cada região.


![]()
REFERÊNCIAS
Cosgrove, A. S. An apparently new disease of chickens: avian nephrosis. Avian diseases, 6(3), 385-389, 1962.
Benton, W. J., M. S. Cover, J. K. Rosenberger, and R. S. Lake. Physiochemical properties of the infectious bursal agent (IRA). Avian Diseases. 11:438–445, 1967.
Di Fabio, J., Castro, A. G. D., Gardin, A. Y., Rossini, L. I., Toquin, L. I. D., & Eterradossi, N. Very virulent IBD spreads to South America. World Poultry, 15(9), 1999.
Gimeno, I. M. and Schat, K.A. Virus-Induced Immunosuppression in Chickens. Avian DiseaseS 62:272–285, 2018.
Arafat, N., Eladl, A. H., Mahgoub, H., El-shafei, R. A. Effect of infectious bursal disease (IBD) vaccine on
Salmonella Enteritidis infected chickens. Vaccine, 35, 3682–3689, 2017.
Hirai, K., Calnek, B.W. In vitro replication of infectious bursal disease virus in established lymphoid cell lines and chicken B lymphocytes. Infect. Immun. 25 (3), 964–970, 1979.
Hirai, K., Funakoshi, T. Sequential changes in the number of surface immunoglobulin- bearing B lymphocytes in infectious bursal disease virus infected chickens. Avian Diseases, 25 (2), 484–496, 1981.
Rodenberg, J., Sharma, J.M., et al. Flow cytometric analysis of B cell and T cell subpopulations in specificpathogen-free chickens infected with infectious bursal disease virus. Avian Diseases, 38 (1), 16–21, 1994.
C. E. Whitfill, E. E. Haddad, C. A. Ricks, J. K. Skeeles, L. A. Newberry, J. N.Beasley, P. D. Andrews, J. A. Thoma and P. S. Wakenell. Determination of Optimum Formulation of a Novel Infectious Bursal Disease Virus (IBDV)
Vaccine Constructed by Mixing Bursal Disease Antibody with IBDV. Avian Diseases, 39, 4, 687-699, 1995.
E. E. Haddad, C. E. Whitfill, A. P. Avakian, C. A. Ricks, P. D. Andrews, J. A. Thomaand P. S. Wakenell. Efficacy of a Novel Infectious Bursal Disease Virus Immune Complex Vaccine in Broiler Chickens. Avian Diseases, 41, 4,882-889, 1997.
Cazaban, C., Swart, W.B.F. Rietema RMW, Wit JJD, Palya V, et al. Field Assessment of An Immune-Complex Infectious Bursal Disease Vaccine in Chicks Born to Non- Hyperimmunized Broiler Breeders. J Vet Sci Ani Husb, 6(3): 302, 2018.
Sedeik, M.E., El-shall, N. A., Awad, A.M., El-Hack, M. E. A., Alowaimer, A. N., Swelum,
A. Comparative Evaluation of HVT-IBD Vector, Immune Complex, and Live IBD Vaccines against vvIBDV in Commercial Broiler Chickens with High Maternally Derived Antibodies. Animals, 9, 72, 2019.
Jeurissen, S.H., Janse, E.M., Lehrbach, P.R., Haddad, E.E., Avakian, A., Whitfill, C.E. The working mechanism of an immune complex vaccine that protects chickens against infectious bursal disease. Immunology 95,494–500, 1998.
Prandini, F, M Bublot, FX Le Gros, A Dancer, L Pizzoni, et al., 2008. Assessment of the immune response in broilers and pullets using two ELISA kits after in ovo or day-old vaccination with a vectored HVT + IBD vaccine (VAXXITEK® HVT+IBD). Zootecnica International, Sept2008: 40-50.
Bublot, M, N Pritchard, FX Le Gros and S Goutebroze. Use of a vectored vaccine against infectious bursal disease of chickens in the face of high-titred maternally derived antibody. J Comp Pathol, 137 Suppl 1:S81-84, 2007.
Bublot, M. Vector vaccines for poultry: their advantages and limitations compared to classical vaccines.
Proceedings of the “International Seminar on Poultry Diseases” 14- 15 Dec, p.7-17, 2015.
European Medicines Agency Veterinary Medicines. EUROPEAN PUBLIC ASSESSMENT REPORT (EPAR).
VAXXITEK HVT+IBD, EPAR. Summary for the public. EMEA/V/C/065.
European Medicines Agency Veterinary Medicines. EUROPEAN PUBLIC ASSESSMENT REPORT (EPAR). InnovaxND-IBD, EPAR. Summary for the public. EMA/391280/2017 – EMEA/V/C/004422.
Phillips, R. A; Opitz, H. M. Pathogenicity and persistence of Salmonella enteritidis and egg contamination in normal and infectious bursal disease virus- infected leghorn chicks. Avian Dis, 39:778–87, 1995.
Sheela, R. R; Babu, U; Mu, J; Elankumaran, S; Bautista, D. A; Raybourne, R. et al. Immune responses against Salmonella enterica serovar enteritidis infection in virally immunosuppressed chickens. Clin Diagn Lab Immunol, 10 (4):670–9, 2003.
Wyeth, P. J. Effect of infectious bursal disease on the response of chickens to S. typhimurium and E. coli infections. Vet Rec, 96:238–43, 1975.


www.boehringer-ingelheim.com.br
PDF