



Descrita na Hungria em 1907 pelo médico veterinário Jozsef Marek, a doença de Marek (DM) adquiriu grande importância durante o século XX com o crescimento da avicultura. Após a descrição da etiologia viral em 1967- 1968, foram desenvolvidas com sucesso as estratégias preventivas de DM baseadas em vacinação, que representaram em amplo controle da doença.
Os estudos da DM em galinhas permitiram avanço no conhecimento das doenças tumorais em animais e têm servido de modelo para os estudos de alguns tipos de câncer em humanos [13]. A DM foi a primeira doença tumoral prevenida por vacinação na história da medicina.
Outro aspecto médico importante é a alteração arterial (arterioesclerose), forma não neoplásica da doença, sendo hoje modelo para as arterioescleroses causadas por vírus. A DM causou, por muitos anos, grande impacto em avicultura de galinhas, e seus efeitos negativos foram reduzidos de forma espetacular, como se “retirados com a mão”, após o uso das vacinas em pintinhos no primeiro dia de idade e, mais recentemente, de forma alternativa, em embriões aos 18 dias de incubação.
Atualmente são raros os episódios da DM e muitos estão relacionados a falhas vacinais, por falta de título vacinal ou doença imunodepressora que impeça a formação de resposta imune protetora, ou ainda, variação de vírus de campo [12]. A vacinação embrionária contra a DM representa antecipação vacinal, que tem como objetivo reduzir a probabilidade de infecção com estirpes de alta virulência emergentes. Além de galinhas, DM natural e experimental foi descrita em codornas japonesas e em perus, e a reprodução experimental em faisões [17].
Etiologia
A DM é causada por vírus denominado GaHV-2 – Marek’s disease virus ou GaHV-2 – Gallid herpesvirus 2, da família Herpesviridae, subfamília Alphaherpesvirinae e gênero Mardivirus [9, 14].
O GaHV-2 é classificado em três grupos distintos, denominados como sorotipo 1, 2 e 3. O sorotipo 1 contém todas as estirpes patogênicas, enquanto as estirpes do sorotipo 2 não são patogênicas. Integrantes do sorotipo 2 são representados pelos isolados de perus, não patogênicos e assemelhados antigenicamente a GaHV-2, e por isso, utilizáveis em vacinas (HVT, herpesvírus de peru) [3, 12, 17].
O GaHV-2, como todos os herpesvírus, contém genoma de DNA de fita dupla muito longa (125.000 pares de bases), 30 proteínas estruturais confirmadas e 10 propostas, com 11 presentes no envelope, sendo destas, pelo menos 10 glicosiladas [9, 14]. Ainda, como todos os herpesvírus, o GaHV- 2 promove infecção persistente e vitalícia (latência), com ciclos de reativação modulados pela resposta imune [14]. As estirpes integrantes do sorotipo 1 (patogênico) diferem em patogenicidade [3, 16]. Nos anos 1980 houve o surgimento de variantes de alta virulência (vvMDV-very virulent Marek’s disease virus), possivelmente por pressão seletiva exercida pela imunidade dos plantéis [3, 5, 12, 17]. Na pele descamada presente na poeira dos galpões, o GaHV-2 está protegido e muito estável, sendo a desinfecção ineficiente no ambiente de galpão, apesar de GaHV-2 extracelular ser muito sensível aos detergentes e desinfetantes [2, 7, 12, 17].
Transmissão e patogenia
Galinhas portadoras transmitem via horizontal vírus para pintinhos sensíveis, por pele descamada eliminada pela ave portadora, que contém vírus infectante [2, 7]. As infecções por GaHV-2 têm alta disseminação nas regiões avícolas [12, 17]. As galinhas de criação industrial e de subsistência portadoras de GaHV-2, inclusive as aves vacinadas, eliminam e transmitem vírus de forma cíclica.
A DM é bastante comum na avicultura de subsistência em Minas Gerais (Martins et al., dados não publicados). Essas aves podem atuar de forma significativa como reservatório de vírus patogênicos. Apesar de haver vacinação obrigatória em 100% da avicultura comercial, pode ocorrer infecção (desafio) natural com vírus de campo [11] em aves vacinadas, embora nenhuma ave vacinada desenvolva tumores, por haver proteção antitumoral. Assim, aves (galinhas) vacinadas podem ser fonte de vírus patogênico, apesar de protegidas contra tumores [11], razão para serem mantidas estratégias de biosseguridade entre plantéis.
O GaHV- 2 é transmitido pelo contato indireto ou direto entre as aves. O pintinho inala poeira do ambiente de criação, que contém pele descamada contendo vírus infectante [2, 7]. Nos capilares aéreos pulmonares, a poeira é fagocitada pelos macrófagos e ocorre a infecção destes. O vírus livre e/ou os macrófagos infectados entram em circulação e transferem infecção para os linfócitos B no baço e na bolsa cloacal (de Fabricius), que resulta em grave destruição de linfócitos B nesses órgãos e imunodepressão.
Na fase inicial, a replicação e a lise de linfócitos se estendem para os linfócitos T do timo. Nessa fase da infecção, a atrofia do timo, do baço e da bursa pode ser identificada pela macroscopia e histologia desses órgãos (17). A viremia persiste e permite que, nos linfócitos T (do timo), ocorra a integração do genoma de GaHV-2 no genoma celular, resultando em transformação dos linfócitos T ativos em linfócitos T neoplásicos.


Ave de subsistência apresentando a forma
clássica da doença de Marek, caracterizada por
paralisia unilateral resultante da infiltração dos
linfoblastos T transformados e por processo inflamatório
nos nervos. Um vídeo da mesma imagem
pode ser acessado em https://www.youtube.com/watch?v=BydP994Tp7E.
A forma clássica de apresentação clínica é caracterizada por paralisias tipicamente unilaterais (Fig. 1), decorrentes da infiltração dos linfócitos T transformados nos nervos, com intenso processo inflamatório e de progressão crônica com letalidade moderada (até 30%) [12, 17, 18].
A forma aguda é de emergência mais rápida e mais grave, com impacto de morbidade e mortalidade altas, que resultam de infiltração tumoral disseminada para órgãos vitais [12, 17, 18].
A patogenia de GaHV-2 está principalmente determinada pela habilidade em transformar linfoblastos T em tumorais [12, 17]. Os linfócitos T transformados colonizam diversos tecidos (metástases).
Na forma clássica, as alterações clínicas típicas ocorrem após as infiltrações linfoblásticas em nervos periféricos (causando perda e degeneração axonal), principalmente os de maior calibre, como os nervos ciático (Fig. 2), braquial, vago e intercostais.
Os plexos dos nervos braquiais e ciáticos também são frequentemente acometidos [12, 17]. As paralisias de MD no membro posterior podem resultar na posição de perna estendida, tipicamente unilateral, por infiltrações tumorais no nervo ciático. Podem também estar paralisadas as asas, os músculos intercostais, a região lombar e os órgãos inervados pelo vago. A forma paralítica ou clássica resulta em processo crônico de emagrecimento e fraqueza, em parte resultantes da dificuldade de
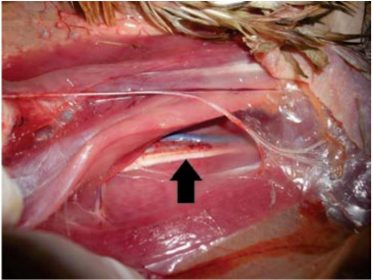
. Ave doença de Marek mostrada na Fig. 1.
Nervo ciático com espessamento (seta) leve decorrente
da infiltração de linfoblastos ativados e
T transformados.
locomoção para alimentação.
A forma aguda da DM resulta em mortalidade mais grave e rápida e está associada ao desenvolvimento de tumores que impedem a função de órgãos vitais, como o fígado, os rins e o baço. Os tumores podem afetar a fertilidade (ovários, testículos) e atingir também o olho, na íris (causando fechamento da pupila) e músculos esqueléticos.
A forma visceral pode ocorrer com alterações macroscópicas neurais pouco perceptíveis ou ausentes. Quando há envolvimento do sistema nervoso central (cérebro e cerebelo), linfócitos inflamatórios se infiltram ao redor dos vasos. Posteriormente, são substituídos por linfoblastos neoplásicos que invadem também o parenquima adjacente.
A forma cutânea é caracterizada por infiltrações de linfoblastos nos folículos das penas. Quando a proliferação é intensa, pode invadir a epiderme e causar ulcerações [17], que, presentes na pele da região da cabeça (periocular), podem ser confundidas com bouba aviária, sendo importante a histopatologia para o diagnóstico diferencial.
Diagnóstico
A forma clássica de DM pode ser suspeita em aves, especialmente galinhas, com paralisia de pernas (mais comum unilateral) (Fig. 1), asas, pescoço e outros, especialmente em aves não vacinadas. As alterações nos nervos podem ser caracterizadas por aumento localizado ou simétrico no diâmetro do nervo, com hiperemia e/ou hemorragia (Fig. 2).
Estes são quadros comuns em galinhas de fundo de quintal ou de subsistência. Em falhas vacinais ou depressão imune, em frangos de corte ou frangas pré-postura, as lesões de pele, com folículos das penas aumentados e/ou hemorrágicos, podem ser sugestivas, especialmente se encontrados tumores em vísceras.
A confirmação do diagnóstico depende de visualização microscópica das células tumorais e/ ou detecção do antígeno [12, 17, 18] ou genoma viral nos tecidos [1, 3, 12, 17].
Os tecidos acometidos devem ser enviados para histopatologia, para a observação de infiltrados de células linfoblásticas pleomórficas (diagnóstico diferencial de leucose linfoide, com células linfoblásticas monomórficas). Na forma inflamatória, pequenos linfócitos e plasmócitos também estão presentes.
Por imuno-histoquímica, linfócitos T podem ser demonstrados nos tumores com anticorpos mono ou policlonais específicos para linfoblastos T, ou GaHV-2 pode ser detectado nos tecidos infectados, usando anticorpos dirigidos para antígenos virais [12, 17, 18]. Na doença de Marek, os tumores são de origem linfoblástica T e diferentes de linfoblastos B presentes na leucose linfoide [18].
Reações em cadeia pela polimerase (PCR) estão descritas para a amplificação de segmentos de genes que codificam proteínas estruturais e não estruturais de GaHV-2 [1, 3, 8, 12, 17, 20, 22]. PCR com oligonucleotídeos específicos, por exemplo, dirigidos para a amplificação dos genes que codificam as glicoproteínas B, C, D, timidina quinase (TK) ou a proteína da célula infectada (ICP-4 – infected cell protein 4) [1, 3, 12, 17], permitem a confirmação da presença viral.
O GaHV-2 pode ser isolado dos tecidos infectados em ovos embrionados (membrana corioalantoide) [19] e em monocamadas primárias ou de linhagens contínuas [18]. Os isolados podem ser classificados quanto ao patotipo, uma vez que há diferenças em patogenicidade entre estirpes do sorotipo 1 [12, 17, 21].
Prevenção e controle
A vacinação contra a DM é obrigatória para todas as galinhas e frangos industriais no Brasil [4] e tem resultado em controle muito eficiente da ocorrência da doença. A vacinação in ovo é feita no 18º dia de incubação (ou seja, antes da eclosão), e a vacinação do primeiro dia de vida (ainda no incubatório) é feita via subcutânea.
As vacinas contra a DM utilizadas no Brasil são sempre vivas, simples ou combinadas, e com um mínimo de 1.500 unidades formadoras de placas. Há vacinas preparadas com estirpes do GaHV-2 sorotipo 1 atenuadas (por exemplo, CVI988/Rispens), sorotipo 2 (por exemplo, SB1) naturalmente não patogênicas, ou vacina do sorotipo 3, herpesvírus de perus (HVT, herpesvirus of turkeys).
As vacinas com HVT são as mais comumente utilizadas, preparadas com a estirpe FC126, relacionada antigenicamente ao GaHV-2, e podem ser conservadas liofilizadas (sublimação a -40°C) ou em nitrogênio líquido (vírus associado à célula). Todas as vacinas com estirpes de GaHV-2 são associadas à célula e mantidas em nitrogênio líquido [12, 17].
Referências
1. Barrow, A. and K. Venugopal. 1999. Molecular characteristics of very virulent European GaHV-2 isolates. Acta Virologica, vol. 43(2-3):90-93.
2. Beasley, J. N., L. T. Patterson, and D. H. McWade. 1970. Transmission of Marek’s disease by poultry house dust and chicken dander. American Journal of Veterinary Research 31:339-344.
3. Becker, Y., Y. Asher, E. Tabor, I. Davidson, M. Malkinson, and Y. Weisman. 1992. Polymerase chain reaction for differentiation between pathogenic and non-pathogenic serotype 1 Marek’s disease viruses (GaHV-2) and vaccine viruses of GaHV-2-serotypes 2 and 3. Journal of Virological Methods 40:307-322.
4. Brasil. MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO, GABINETE DO MINISTRO, INSTRUÇÃO NORMATIVA Nº 56, DE 4 DE DEZEMBRO DE 2007.
5. Buscaglia, C., Nervi, P., & Risso, M. 2004. Characterization of four very virulent Argentinian strains of Marek’s disease virus and the influence of one of those isolates on synergism between Marek’s disease vaccine viruses, Avian Pathology, 33:2, 190-195.
6. Calnek, B. W. and S. B. Hitchner. 1969. Localization of viral antigen in chickens infected with Marek’s disease herpesvirus. Journal of the National Cancer Institute 43: 935-949.
7. Calnek, B. W., A. M. Alexander, and D. E. Kahn. 1970. Feather follicle epithelium: a source of enveloped and infectious cell-free herpesvirus from Marek’s disease. Avian Dis. 14: 219-233.
8. Davidson, I., A. Borovskaya, S. Perl, and M. Malkinson. 1995. Use of the polymerase chain reaction for the diagnosis of natural infection of chickens and turkeys with Marek’s disease virus and reticuloendotheliosis virus. Avian Pathol 24:69-94.
9. Index of Viruses – Herpesviridae (2006). In: ICTVdB – The Universal Virus Database, version 4. Büchen-Osmond, C (Ed), Columbia University, New York, USA. http://www.ncbi.nlm.nih.gov/ ICTVdb/Ictv/fs_index.htm
Cadernos Técnicos de Veterinária e Zootecnia, nº 76 – março de 2015