



Conteúdo disponível em:
A coccidiose aviária é uma doença parasitária causada por protozoários apicomplexos do gênero Eimeria. Sua importância na indústria avícola moderna é tal, que sem adequadas medidas de controle – quimioterapia e vacinação -, o setor não teria chegado à sua atual condição. Neste trabalho serão brevemente revisadas a etiologia e patogenia das espécies de Eimeria que parasitam a galinha poedeira ou reprodutora e o frango de corte.
Espécies de Eimeria que parasitam as aves de produção
Nas aves de produção – Gallus domesticus – foram relatadas até 15 espécies diferentes. No entanto, os critérios a serem considerados para o reconhecimento de uma espécie foram se desenvolvendo progressivamente, sempre posteriormente às descrições originais das mesma, razão pela qual hoje se questiona a validade taxonómica de algumas delas.

Os métodos tradicionais para a identificação específica incluem:
- Estudo de várias características fenotípicas – morfológicas e biológicas -, como tamanho, forma e tempo de esporulação dos oóscistos.
- Localização do parasita na mucosa.
- Lesões que produz.
- Região principal de parasitação.
- Desenvolvimento endógeno – número de esquizogonias, tamanho dos esquizontes, número de merozoitos que contém e seu tamanho etc.
- Imunização cruzada com outras espécies e diferenças em sua composição enzimática.
Estas técnicas, ainda que onerosas em tempo e dinheiro, são necessárias, complementando-se com os atuais métodos moleculares que incluem técnicas de PCR baseadas na amplificação do DNA. Além disso, a já próxima sequenciação do genoma de todas as espécies de Eimeria que parasitam as aves de produção permitirá no futuro um grande avanço neste e outros campos da coccidiose aviária.
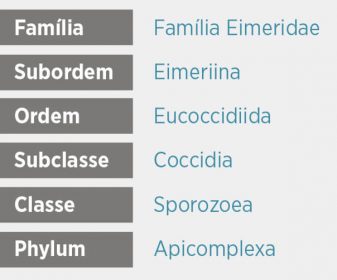
Atualmente consideramos sete espécies de Eimeria que parasitam a Gallus domesticus

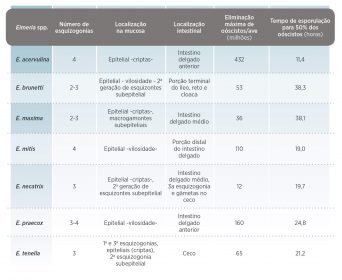
Tabela 1. Espécies de Eimeria que parasitam as aves de produção. São indicadas algumas diferenças de seu ciclo biológico.
Ciclo biológico das Eimeria spp. aviárias
As sete espécies de Eimeria têm um ciclo biológico similar, sendo todas parasitas intracelulares intestinais.


O ciclo biológico compreende duas fases de reprodução endógena no intestino da ave:
- Uma assexual – esquizogonia;
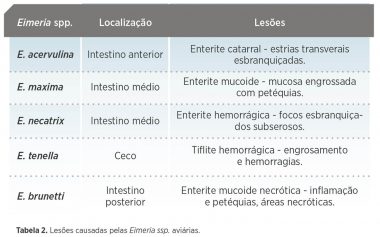
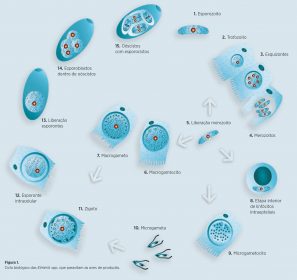
- Outra sexual – gamogonia – seguidas de outra fase assexual exógena – esporogonia no ambiente que termina com a formação de esporozoitos no oóscisto – figura 1.


TRANSMISSÃO
A transmissão é direta entre as aves mediante a ingestão dos oóscistos esporulados que se encontram no ambiente contaminando, água e alimento. A infecção é autolimitante na ausência de reinfecções.


Fase exógena ou ambiental do ciclo biológico
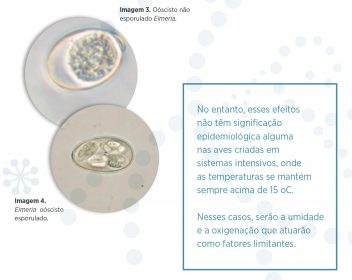
Os oócistos não esporulados são o estado exógeno que aparece nas fezes do hospedeiro – Imagem 3. Estes oócistos não esporulados carregam, sob uma dupla membrana, uma massa citoplasmática não diferenciada chamada esporonte.
O processo de esporulação inicia quando as condições de aeração, temperatura e umidade são adequadas para isso. O esporonte, por divisão nuclear dá lugar a quatro massas ovaladas – esporoblastos – que se recobrem de uma dupla membrana, formando, assim, quatro esporocistos, cada um dos quais contendo dois esporozoitos. Estes esporozoitos são as formas infectantes para as aves – Imagem 4.

INFLUÊNCIA DA TEMPERATURA NA ESPORULAÇÃO
A esporulação não acontece abaixo de 12ºC, nem acima de 39ºC, sendo o ideal de temperatura entre 28 – 31ºC.
- A inibição da esporogonia por exposição dos oóscistos a baixas temperaturas costuma ser temporal, permitindo a esporulação posterior, quando a temperatura passa de 12ºC.
- A exposição prolongada a baixas temperaturas -p.e. 26 semanas a 4ºC destroi sua viabilidade e foi comprovado que o congelamento de oóscistos não esporulados de E. tenella durante 20 dias impede sua esporulação, assim como o congelamento e descongelamento diários.
Os tempos de esporulação para cada uma das diferentes espécies que infectam as aves de produção são indicados na -Tabela 1 p.23-. A viabilidade dos oóscistos uma vez esporulados é limitada, influenciando de maneira decisiva o conteúdo em carboidratos – amilopectinas – dos esporozoitos. Estas reservas diminuem não só com a idade, como também de forma inversamente proporcional à temperatura de armazenamento.
Fase endógena do ciclo biológico
Quando a ave ingere os oóscistos esporulados, os esporozoitos devem ser liberados de suas paredes protetoras e parasitar a célula intestinal. Figura 1 p.25, Ciclo biológico das Eimeria spp. que parasitam as aves de produção. Esse processo de desencistamento exige dois estímulos separados:
- A ação mecânica da moela sobre os oóscistos que alteraria a membrana permitindo a passagem de sais biliares.
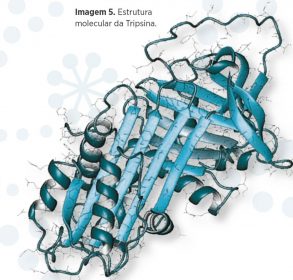
- A ação dos sais biliares que facilitariam a entrada de enzimas, como a tripsina ou pancreatina, alterando a superfície do corpo da Stieda e permitindo a saída dos esporozoitos do esporocisto a partir de movimentos ativos – nos quais a bilis pode atuar como estimulante – ver imagem 5.

REQUISITOS DE BILIS
- Os requisitos de bilis variam segundo as espécies, sendo a E. tenella menos dependente que outras Eimeria spp.
- Observou-se que a maioria dos esporocistos de E. tenella encontravam-se no ceco uma hora depois da inoculação experimental de oóscistos via oral e que o desencistamento dos mesmos deveria ocorrer no ceco.
- Essa baixa receptibilidade da E. tenella à ação biliar atrasa o desencistamento e poderia ser um fator chave na localização cecal deste parasita.
A IDADE DO HOSPEDEIRO
- A idade do hospedeiro no momento da infecção também parece influir no desencistamento dos oóscistos, sendo a taxa de desencistamento menor em pintos de um dia, do que nos de três dias.
- Possivelmente, isso seria causado pela menor capacidade da moela para romper os oóscistos e a quantidade de enzima pancreática segregada.
INVASÃO DOS ENTERÓCITOS SEGUNDO A ESPÉCIE DE EIMERIA
- Os esporozoitos invadem os enterócitos superficiais e são transportados através de linfócitos intraepiteliais, dirigindo-se através da lâmina própria às criptas de Lieberkün. Esta migração ocorre na E. acervulina, E. maxima e E. tenella.
- Na E. brunetti e E. praecox, nas quais não ocorre traslocação de esporozoitos do epitélio às criptas, também foram encontrados esporozoitos em linfócitos intraepiteliais.
- Inclusive no caso da E. mitis, que parasita a base da vilosidade ou as criptas, foram encontrados esporozoitos em leucócitos intraepiteliais nas criptas.
- Os esporozoitos de E. necatrix invadem diretamente as criptas de Lieberkün, não sendo nunca fagocitados por macrófagos, nem realizado o transporte por outras células.
MIGRAÇÃO DOS ESPOROZOITOS
Durante estas migrações os esporozoitos podem penetrar na circulação sanguínea e chegar a outros tecidos, conseguindo transferir a infecção, a partir do sangue ou de extractos de tecido – baço e fígado -, de E. tenella e E. acervulina ou de E. acervulina, E. brunetti, E. maxima, E. necatrix e E. praecox, respectivamente. Estes feitos poderiam ser interpretados da seguinte forma: a maioria dos esporozoitos deixava o intestino para passar via sanguínea a outros órgãos e, dali, liberar-se de maneira mais lenta.
Uma vez invadida a célula, o esporozoito se arredonda no interior transformando-se em trofozoito, que por divisão nuclear reiterada – esquizogonia – origina um esquizonte polinuclear de primeira geração.
No entanto, ao menos na E. tenella essa migração é muito escassa – a inoculação de esporozoitos em uma das duas pontas do ceco previamente ligada produz oóscistos quase exclusivamente no ceco inoculado, enquanto que a inoculação via intravenosa superficial produz infecções de intensidade similar em ambas as pontas do ceco.

ESPÉCIES DE EIMERIA
- E. brunetti, E. necatrix e E. tenella, a segunda geração de esquizontes é de localização subepitelial, porém parasitando exclusivamente células epiteliais.
- E. tenella e E. necatrix, as mudanças na célula infectada estimulam sua migração à lâmina própria, rompendo a membrana basal da cripta.
- Nas espécies mais patógenas, E. necatrix e E. tenella, existe uma terceira esquizogonia.
GAMETAGONIA
Finalizadas as esquizogonias, os merozoitos penetram em novas células para diferenciar-se em gametócitos.
Ainda que o mecanismo de diferenciação sexual não esteja o suficientemente claro, parece haver um merozoito determinado masculino ou femininamente, em virtude do número de grânulos de polissacarídeos ou de mitocondrias observadas, o desenvolvimento do retículo endoplasmático e a presença ou não de vacúolos nutricias.
O merozoito de determinação masculina evolui para microgamonte e, após uma fase de multiplicação de seu núcleo – mitose – e outra de estrangulamento, dá lugar a múltiplos microgametas flagelados.
O merozoito de determinação feminina evolui para macrogamonte e em seguida para macrogameta.
Os microgametas livres invadem as células parasitadas pelos macrogametas fecundando-os e dando lugar ao zigoto. A fusão dos corpos de revestimento I e II do macrogameta dá lugar às capas externas do oóscisto no esporulado. Ao romper-se a célula que o alberga, o oóscisto alcança a luz intestinal ou cecal e, posteriormente, o ambiente através das fezes – Figura 1.
Especificidade da Eimeria spp.
ESPECIFICIDADE
Hospedadeiro: as espécies de Eimeria apresentam um marcado estenoxenismo, ou seja, é raro que o parasita complete seu ciclo em mais de um gênero de ave.
Imunológica: a resistência à reinfecção obtida com uma espécie não protege contra outras.
Localização: é também muito delimitado pela espécie e lugar de parasitação. No entanto, pode-se fazer algumas exceções ou matizações.
Foram isoladas espécies de coccídios em galos selvagens – Gallus lafalletique infectavam o doméstico – Gallus domesticus -, espécies que, após estudos de transmissão cruzada foram consideradas como E. praecox var. celonensis, E. maxima var. identata e E. acervulina var. diminuta. Também se reproduziu E. tenella isolada de G. domesticus na perdiz grega – Alectoris graeca – e infectou-se híbridos frangos/codornas com oóscistos de frangos – E. acervulina, E. maxima e E. tenella – de codornas -Eimeria bateri -, detectando alguns oóscistos após o período de patência normal.


Imagem 7. Gallus gallus
-Gallus domesticus-.
Fora esses casos, não há evidências que demonstrem a transmissão completa de outras espécies, nem sequer entre espécies de galináceos afins, ainda que se tenha conseguido infectar frangos com Eimeria meleagrimitis do peru -Meleagris gallopavo.


Imagem 8. Perdiz grega -Alectoris graeca.
Cada espécie de Eimeria tem um lugar de preferência para seu estabelecimento no intestino, parasitando quase exclusivamente células epiteliais -Tabelas 1 e 2.
No entanto, algumas delas podem completar seu ciclo em tecidos diferentes dos habituais, como o tecido cecal e embrião do frango, onde outras não são capazes de fazê-lo. Determinados estados do ciclo, em algumas espécies de Eimeria, podem migrar a certos órgãos como fígado e baço.
Não se conseguiu que a infecção com uma espécie de Eimeria ofereça imunidade contra outra diferente, no sentido de que proporcione completa proteção ou proteção similar à inmunidade conferida contra seu próprio desafio. No entanto, deve-se destacar a heterogeneidade imunológica entre cepas de uma mesma espécie. Como o caso da E. acervulina, E. mitis, E. maxima, E. necatrix e E. tenella. As diferenças imunológicas mais importantes foram observadas na E. maxima. Pôde-se comprovar que o cultivo misto de cepas de E. maxima confere proteção contra o desafio heterólogo de isolados de campo da mesma espécie.
Experimentalmente, a E. tenella completa seu ciclo em tecidos adjacentes ao ceco quando este foi cirurgicamente extirpado e sua especificidade de localização pode ser reduzida mediante tratamento com corticosteroides, podendo realizar a esquizogonia no fígado de frangos tratados com dexametasona.
Nas outras Eimeria spp. as diferenças de proteção verificadas não foram tão chamativas. Esse aspecto é de grande importância no desenvolvimento de vacinas para uso industrial na produção avícola. As variações imunológicas entre cepas de uma mesma especie poderiam repercutir no funcionamento destas vacinas no campo.
Patogenia
Infecção intestinal
A afecção intestinal causada por qualquer das espécies de Eimeria segue um caminho mais ou menos grave, determinado por um grande número de variáveis como
- Idade.
- Estado sanitário e imunitário das aves no momento da infecção.
- Número de oóscistos ingeridos.
- Espécies de Eimeria implicadas etc.
A patogenicidade intrínseca das Eimeria spp. parece estar diretamente relacionada com o lugar de desenvolvimento, de maneira que as espécies mais patógenas são as que penetram mais diretamente na mucosa, sem esquecer a destruição do tecido epitelial da vilosidade intestinal produzida em todas elas.
ESPÉCIES DE EIMERIA RESPONSÁVEIS PELOS QUADROS MAIS GRAVES
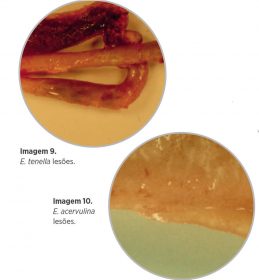
- As mais patógenas são E. tenella e E. necatrix. Ambas espécies produzem hemorragias a partir de finais do 4º e princípios do 5º dia pós-infecção, que se associam à maturação dos esquizontes de segunda geração e produzem uma elevada taxa de mortalidade – Imagem 9.
- E. brunetti e E. maxima causam enterities mucoide, frequentemente com fezes manchadas com sangue e ocasionando, às vezes, mortalidade e quadros de coccidiose muito graves.
- E. acervulina se associa com enterite não hemorrágica -aquosa- e produz, principalmente, diarreias aquosas que originam perda no ganho de peso -Imagem 10.
- As outras duas espécies de Eimeria, E. mitis e E. praecox, não produzem mortalidade, nem lesões manifestamente visíveis, associando-se sua infecção a uma redução do ganho de peso e observando-se diminuição na absorção intestinal durante a infecção

A PATOGENICIDADE DE UMA ESPÉCIE DE EIMERIA VARIA CONFORME A CEPA
Diferentes isolados de uma mesma espécie apresentam diferente patogenicidade intrínseca.
A interação entre espécies de coccídios pode se desenvolver seguindo caminhos diferentes, assim as espécies que parasitam a mesma região intestinal – como E. mitis e E. brunetti – competem e o efeito combinado não é maior que quando a infecção é produzida por uma só espécie. Porém, entre espécies que parasitam diferente região intestinal – como E. mitis e E. acervulina – o efeito patógeno da combinação é muito maior.
Outros fatores que não estão diretamente relacionados com a patogenicidade intrínseca das espécies de Eimeria podem afetar o resultado final da infecção, como a idade e genética das aves ou as interações com outras patologias. Assim, pôde-se comprovar que os animais adultos que foram mantidos livres de coccídios durante sua crianção são mais receptivos à infecção que os jovens. Nestes existe uma maior reprodução do parasita, os sinais clínicos e a mortalidade são mais importantes; e o ganho de peso é menor. Também foram descritas diferenças genéticas observadas na resistência à coccidiose entre diferentes raças ou estirpes de aves.
Em infecções com uma mesma cepa, sua patogenicidade está diretamente relacionada com a quantidade de oóscistos que ingere a ave, ainda que esta relação seja limitada, de maneira que acima de um nível de infecção, doses maiores não conferem maiores lesões nem produzem mais quantidade de oóscistos.
Um aparte importante é a interação com outras patologias, como as deficiências nutricionais ou as infecções causadas por bactérias, vírus, parasitas e micotoxinas. Essas interações são cada vez mais importantes, conforme os avanços da genética e nutrição produzem aves de crescimento mais rápido.
Foram realizados inúmeros estudos sobre os efeitos de diferentes componentes da dieta, como a L-arginina, treonina, diferentes vitaminas e produtos naturais constituídos por diferentes plantas sobre a coccidiose aviária. Pôde-se demonstrar a redução da absorção de glicose, ácido oleico, vitamina A, carotenoides, cálcio e elementos traço, como o zinco, no duodeno de aves infectadas com E. acervulina. Os efeitos com outras espécies são desconhecidos em grande medida, ainda que deve-se considerar que no caso da glicose e aminoácidos, por exemplo, a redução da absorção duodenal pode ser compensada com um aumento da mesma no íleo.
A infecção por Eimeria spp. também pode ocasionar mudanças na permeabilidade da mucosa, resultando na perda de proteínas plasmáticas à luz intestinal, acompanhada do aumento do pH e diminuição da motilidade.
INFECÇÕES CONJUNTAS
Infecções com E. maxima são acompanhadas de uma marcada hipoproteinemia e redução das concentrações plasmáticas dos hormônios tiroxina e triiodotironina, asim como um aumento da corticosterona e prolactina. No entanto, não se observou mudanças no hormônio do crescimento, indicando que este não está envolvido na diminuição do crescimento observado durante a coccidiose.
Ainda que a patologia intrínseca da E. tenella seja similar nos frangos gnotobióticos e convencionais, as infecções conjuntas de E. tenella e bactérias em frangos monoflora são mais graves que as infecções de coccídios sozinhas. Assim, a capacidade de invasão da Salmonella tiphimurium – Imagem 12 – é favorecida em infecções concorrentes com E. necatrix, E. acervulina e E. maxima; assim como E. tenella, respectivamente. O mesmo ocorre em infecções concorrentes de Clostridium perfringens – Imagem 11 – com E. tenella, E. acervulina, E. necatrix e E. brunetti, respectivamente.
Foi descrito um aumento de mortalidade em infecções mistas de E. tenella e Escherichia coli, ainda que não se tenha confirmado agravamento da aerossaculite ou pericardite e infecções conjuntas de E. necatrix com E. coli ou Mycoplasma gallisepticum. As enterobacteriáceas Bifidobacterium thermophilum e Lactobacilus acidophilus em infecções conjuntas com E. tenella experimentam maior crescimento no ceco. A mortalidade aumenta em infecções mistas de E. tenella com aflatoxinas ou com vírus da Doença de Marek, virose que impede o desenvolvimento da imunidade estável contra o coccídio. As infecções mistas de coccídios com vírus da doença de Gumboro associam-se à elevada mortalidade por coccidiose e, ainda que não se aprecie bloqueio do desenvolvimento imunitário contra estes, a resposta parece ser menos eficaz. As infecções concorrentes de reovírus com coccídios produzem diminuição significativa no ganho de peso e aumento de enanismo e problemas de patas, inclusive com baixos níveis de exposição a coccídios.
PDF