



Os primeiros relatos de Influenza Aviária (IA) na América Latina e no Caribe foram registrados na década de 1970 no Brasil, onde a presença do vírus foi detectada por sorologia (imunoaglutinação) em aves silvestres locais e migratórias, assim como em aves domésticas2. Infelizmente, nesses relatos não há dados do subtipo nem de sequência genética.
Outra referência de vigilância epidemiológica da IA foi relatada no México nos anos 1980-1981, sem a detecção de anticorpos contra a doença
em lotes de aves comerciais nas regiões de maior produção e densidade avícola do país7.
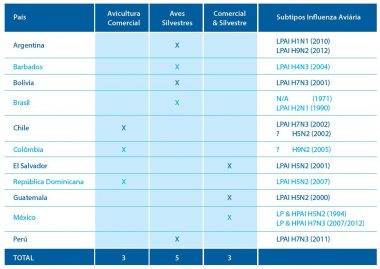
Quadro 1. Resumo epidemiológico da Influenza Aviária na América Latina:

Mais tarde, em 1993 e no início de 1994, os avicultores no México começaram a relatar problemas respiratórios em lotes de frangos de corte, que não eram compatíveis com a doença de Newcastle, coriza, bronquite infecciosa, ou metapneumovírus aviário. Somente em maio de 1994, foram isolados vírus da influenza aviária nos estados de Hidalgo, México e Querétaro, posteriormente confirmados como vírus do subtipo H5N2 de baixa patogenicidade pelo NVSL, pelo laboratório de referência internacional para gripe aviária, localizado em Ames, nos EUA.


O último relato de vírus H5N2 altamente patogênico isolado no México data de 1995. Desde então, todos os isolados de vírus da influenza aviária foram exclusivamente de baixa patogenicidade. Foi também estabelecido um programa de certificação oficial dos estados livres de IA.
No entanto, o vírus de baixa patogenicidade subtipo H5N2 ainda é prevalente em operações avícolas comerciais em certas regiões do México, sendo comumente associado a infecções por outros patógenos aviários, além de representar um desafio e um flagelo constante para a indústria avícola desse país. Na Guatemala (2000) e em El Salvador (2001) foram relatados isolamentos de vírus H5N2 de baixa patogenicidade geneticamente semelhantes ao vírus de IA prevalente no México, além de outros isolamentos esporádicos em ambos os países desde então4. Foi feita vigilância epidemiológica em outros países da América Central (Belize, Costa Rica, Honduras e Nicarágua), sem relatos de infecções de IA.




Supõe-se que a introdução do vírus de baixa patogenicidade subtipo H7N3 tenha ocorrido antes de abril de 2002 e a mutação para alta patogenicidade, provavelmente em um único galpão aviário. Essa mutação de baixa para alta patogenicidade ocorreu rápido. A caracterização do isolamento viral inicial foi feita no final de abril e início de maio de 2002, quando as aves afetadas apresentaram sinais clínicos leves com ligeira queda na postura, sinais clínicos respiratórios e salpingo-peritonite4. A granja de matrizes reprodutoras alojava aves de várias idades distribuídas em 27 galpões. Poucas semanas mais tarde, no final de maio, a gravidade da doença mudou rápido, atingindo mortalidade total acumulada superior a 100.000 aves com forte queda na produção de ovos. Foi observada taxa elevada de mortalidade, intensificando-se sobretudo em alguns galpões, sem apresentação de sinais clínicos nas aves. A queda na postura variou de 10% a 100% nos diferentes galpões.
Considerando-se essa taxa do surto de AI, a doença se espalhou para uma granja próxima de matrizes reprodutoras de perus, com população
de cerca de 50.000 aves distribuídas em 4 galpões sem luz. Para conter o surto, essas duas granjas atingidas com mais de 480.000 aves foram despovoadas. A análise molecular do vírus revelou uma recombinação homóloga entre os genes HA e NP com inserção de 30 nucleotídeos perto do local de ruptura da proteína hemaglutinina (HA), com a presença de três aminoácidos básicos de acordo com a sequência deduzida e com um índice de patogenicidade intravenosa (IPIV) de zero para esse isolado viral de alta patogenicidade, que deveria ser de 2,4 a 3,0 de acordo com os padrões de patogenicidade de IA. O isolado viral chileno revelou-se muito diferente de todos os outros isolados de AI subtipo H7, representando um clade sul-americano totalmente distinto.


em vão4.
Campanhas de vigilância epidemiológica também foram realizadas na região do Caribe entre 2006 e 2009. Essa região é considerada de alto risco para IA por várias razões:
a) Ampla distribuição de aves domésticas;
b) A indústria avícola comercial considerada de risco está presente em várias ilhas (Trinidad, Barbados, Jamaica);
c) Estruturas diferentes dos sistemas de vigilância de doenças aviárias na região;
d) Atividade comercial legal e clandestina com movimentação significativa de aves (Guadalupe e Martinica);
e) A rota migratória de aves da América do Norte e do Sul.




O último caso de IA de alta patogenicidade na América Latina foi relatado em junho de 2012 no México, com a detecção de um vírus subtipo H7N3 no estado de Jalisco, caracterizado por alta densidade populacional de poedeiras comerciais. Esse vírus altamente patogênico se espalhou para outros estados mexicanos, apesar dos esforços para conter a doença com medidas de controle-padrão9.
Conclusões
- Diversos países da América Central, América do Sul e do Caribe fizeram campanhas de vigilância epidemiológica para detecção da influenza aviária em populações avícolas e em aves silvestres (Argentina, Barbados, Belize, Bolívia, Brasil, Chile, Colômbia, Costa Rica, República Dominicana, El Salvador, Guadalupe, Guatemala, Honduras, Martinica, Nicarágua, Peru e Santa Lúcia8).
- Um vírus de IA isolado em aves silvestres na Argentina revelou ter uma linhagem filogenética potencialmente diferente e única para a América do Sul. Além disso, a análise molecular dos isolados dos vírus de IA na Guatemala e no Peru mostrou estreita relação com a linhagem norte-americana.
- Na história da avicultura latino-americana foram relatados três surtos de influenza aviária de alta patogenicidade: um surto do subtipo H5N2 (no México em 1995) e dois surtos do subtipo H7N3 (no Chile em 2002 e no México em 2012). Ademais, dois países na América Central (El Salvador, Guatemala) e um no Caribe (República Dominicana) relataram isolados de baixa patogenicidade subtipo H5N2 em operações avícolas comerciais. Na Colômbia em 2005 houve relato de detecção de anticorpos contra AI subtipo H9N2, sem isolamento viral. Os relatos restantes de isolados do vírus de IA foram em aves silvestres (Argentina, Bolívia, Barbados, Brasil e Peru), sobretudo em patos (Cinnammon teal e Blue-winged teal).
- Por fim, vale destacar o resultado da análise molecular do isolado do vírus altamente patogênico subtipo H7N3 no surto no Chile, apresentando uma recombinação homóloga entre os genes HA e NP com inserção de 30 nucleotídeos perto do local de ruptura da hemaglutinina (HA) contendo três aminoácidos básicos e com um índice de patogenicidade intravenosa (IPI) de zero, que deve ser de 2,4 a 3,0 para os vírus altamente patogênicos.
- Esta breve revisão de casos históricos na detecção e introdução da influenza aviária em diferentes países e regiões da América Latina deixa claro a ameaça dessa doença para operações avícolas comerciais suscetíveis à infecção. Medidas de bioseguridade rigorosas e permanentes (em termos conceituais, estruturais e operacionais) são parte crucial da estratégia de prevenção da doença, para minimizar o risco de introdução da infecção em operações avícolas comerciais. Além disso, as campanhas de vigilância epidemiológica (ativa e passiva) constituem um sistema de alerta permanente que possibilita a detecção precoce e a contenção rápida, para evitar a possível propagação dessa doença altamente contagiosa para grandes populações de aves suscetíveis.


Referências Bibliográficas
1. Buscaglia C. et al.. Avian Influenza Surveillance in Backyard Poultry of Argentina. 2007. Avian Diseases 51: 467-469.
2. Gonzalez-Reiche A. and Perez D. Where Do Avian Influenza Viruses Meet in the Americas? 2012. Avian Diseases 56:1025-1033.
3. Lefrancois T. et al. Surveillance of Avian Influenza in the Caribbean Through the Caribbean Health Network: Surveillance Tools and Epidemiologic Studies. 2010. Avian Diseases 54: 369-373.
4. Senne D.A. Avian Influenza in North and South America, 2002-2005. 2007. Avian Diseases 51:167-173
5. Max V. et al. Avian Influenza in Chile: A successful Experience. 2006. Avian Diseases 51: 363-365.
6. Spackman E. et al. An Avian Influenza Virus from Waterfowl in South America Contains Genes from North American Avian and Equine Lineages. 2007. Avian Disease 51: 273-274.
7. Villareal-C. L. and Flores A. O. The Mexican Avian Influenza (H5N2) Outbreak. 2003. Proceedings Fourth International SYmposium of Avian Influenza. Avian Diseases 47: 18-22.
8. Gherzi B.M. et al. Avian influenza in wild birds in central coast of Peru. 2009. Emerging infectious diseases Vol. 15 No. 6.
9. Bertran K. et al. Protection against H7N3 high pathogenicity avian influenza in chickens immunized with a recombinant fowlpox and an inactivated avian influenza vaccines. 2013. Vaccine 31: 3572–3576
Crédito fotos : Fotolia – S.Klein – O. Langlois – Ceva Santé Animale