


Konten ini tersedia dalam: English ไทย (Thai) Tiếng Việt (Vietnamese) Philipino
Penyakit Gumboro, juga dikenal sebagai Infectious Bursal Disease (IBD) atau Avian Infectious Bursitis, pertama kali dilaporkan di Delaware, AS, pada tahun 1962. Ini adalah penyakit virus imunosupresif yang terutama menyerang ayam berusia antara 3 hingga 6 minggu dan memiliki distribusi global.
- Virus yang menyebabkan penyakit ini termasuk dalam genus Avibirnavirus, famili Birnaviridae, dan memiliki dua serotipe: I dan II.
- Serotipe I telah terdeteksi pada ayam, ayam betina, merpati, dan ayam mutiara (Kasanga et al., 2008) tetapi hanya bersifat patogen pada ayam dan ayam betina.
- Serotipe I selanjutnya dibagi menjadi dua subtipe antigenik: klasik dan varian, sedangkan serotipe II tetap asimtomatik pada kalkun, burung gagak, burung unta, dan bebek (Ogawa dkk., 1998; Yilmaz dkk., 2019).
Sejak laporan pertamanya, banyak varian virus telah teridentifikasi, sehingga menyulitkan upaya pengendalian penyakit. Sampai tahun 1980-an, vaksinasi efektif dalam mengendalikan penyakit, dengan tingkat kematian pada ayam pedaging di bawah 2%.
- Namun, dengan mutasi dan penyusunan kembali virus yang terus berlanjut, varian antigenik baru muncul, yang menyebabkan angka kematian lebih tinggi, bahkan dengan adanya protokol vaksinasi yang ketat.
Varian-varian ini dapat muncul secara subklinis, mengurangi pertumbuhan dan meningkatkan kerentanan terhadap infeksi sekunder, yang mengakibatkan kerugian ekonomi besar bagi industri unggas.
MELIHAT LEBIH JELAS VIRUSNYA
- Virus ini berbentuk icosahedral, tidak memiliki selubung, dan terdiri dari dua segmen RNA untai ganda linear, yang diberi nama A dan B.
Segmen B mengkode VP1, yaitu RNA polimerase virus, sedangkan segmen A menghasilkan protein kapsid pVP2 dan VP3, serta protease VP4 dan VP5, yaitu protein non-struktural yang berperan dalam fungsi regulasi dan disrupsi membran pada sel yang terinfeksi. ( (Mundt, 1999) (Gambar 1).
- Di antara komponen-komponen yang disebutkan di atas, protein VP2 sangat penting karena menentukan antigenisitas, virulensi, dan patogenisitas virus.
Ini berisi wilayah yang dapat diikat oleh antibodi, dan ketika terpapar respon imun, ia cenderung mengalami mutasi yang lebih besar, sehingga menjadikannya wilayah yang sangat bervariasi. (Letzel dan kawan-kawan, 2007).
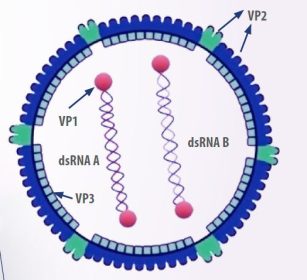

- Protein kapsid VP2 mengandung tiga domain berbeda: base (B), envelope (S), dan projection (P).
- Domain P terdiri dari empat struktur loop yang terekspos pada permukaan virus.
- Loop Pbc (posisi asam amino 219 dan 224) berperan dalam menstabilkan tempat pengenalan antibodi.
- Domain Phi (asam amino 315–324) dikenali oleh antibodi penetral dan merupakan lokasi utama untuk substitusi asam amino yang memungkinkan virus menghindari respons imun.
- Dua loop yang tersisa, Pde (asam amino 250–254) dan Pfg (asam amino 283–287), dikaitkan dengan kemampuan virus untuk menyebabkan penyakit (Jackwood et al., 2018 ( (Gambar 2).

Selain itu, sifat genom virus yang tersegmentasi memudahkan penyusunan ulang genetik antara strain selama koinfeksi. Misalnya, hal ini memungkinkan jenis vaksin live dan virus tipe wild untuk bercampur.
- Sebagai akibat, mutasi dan reassortment berkontribusi terhadap variabilitas antigenik, yang dapat mengurangi efektivitas vaksin komersial terhadap penyakit tersebut, sehingga menimbulkan tantangan yang signifikan dalam pengendalian penyakit. (Gao dan al, 2007).
KEMAJUAN DALAM SKEMA KLASIFIKASI VIRUS
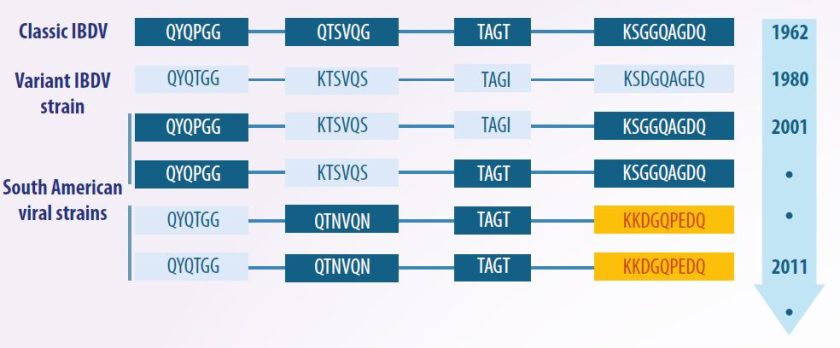
Sejak identifikasi pertamanya pada tahun 1962, strain-strain tersebut belum dikategorikan, karena diasumsikan serupa dalam antigenisitas dan patogenisitas.
- Namun, pada tahun 1980, penemuan serotipe 2 menyebabkan teridentifikasinya varian antigenik, yang disebut sebagai “varian”, serta strain yang sangat virulen, yang dikenal sebagai “strain sangat virulen” (vvIBDV).
Akibatnya, strain yang terdeteksi sebelum varian ini diklasifikasikan sebagai “strain klasik”.
- Metode alternatif digunakan untuk memberi nama strain, berdasarkan nama ilmuwan yang pertama kali mengkarakterisasi isolat (misalnya, Winterfield, Lukert, Moulthrop, Baxendale), lokasi isolat (misalnya, Del-A, Del-E, MD, OH), atau kode alfanumerik (misalnya, STC, D78, G603, S706, 228E).
Namun, seiring berjalannya waktu, karakteristik yang terkait dengan antigenisitas, struktur molekuler, dan patogenisitas kategori ini ditemukan.
- Hal ini mengarah pada skema klasifikasi tradisional, yang membagi strain menjadi klasik dan varian, dengan yang terakhir dibagi lagi menjadi kategori yang dilemahkan, virulen, dan sangat virulen.
Mengenai genom, vvIBDV berbagi residu asam amino spesifik pada posisi 222 (Ala), 256 (Ile), 294 (Ile), dan 299 (Ser) dalam urutan VP2.
Dari segi patogenisitas, dibandingkan dengan strain klasik, vvIBDV cenderung menyebabkan tingkat kematian yang lebih tinggi pada ayam bebas patogen tertentu setelah terinfeksi. (Van Den Berg dan kawan-kawan, 2004).
- Namun, tidak semua strain yang sangat virulen menunjukkan patogenisitas yang tinggi, yang menunjukkan bahwa klasifikasi ini tidak lengkap. ( (Jackwood dan kawan-kawan, 2018).
- Selanjutnya, karena mutasi dan rekombinasi yang berkelanjutan, strain baru tidak dapat diklasifikasikan menggunakan sistem tradisional.
- Hasilnya, Michel dan Jackwood (2017) mengusulkan klasifikasi baru berdasarkan urutan asam amino dari wilayah hipervariabel VP2. Pendekatan ini menghasilkan identifikasi 7 genogrup.
- Akan tetapi, genogrup ini tidak memperhitungkan strain varian baru atau strain yang dilemahkan.
- Lebih jauh lagi, karena rekombinasi karakteristik segmen virus, klasifikasi yang hanya didasarkan pada VP2 tidak sepenuhnya menangkap kompleksitas genogrup virus.


Pada tahun 2021, Wang dkk. mengusulkan sistem klasifikasi baru yang mempertimbangkan karakteristik molekuler VP1 dan VP2, yang masing-masing berasal dari segmen B dan A.
- Pendekatan ini menghasilkan identifikasi 9 genogrup untuk segmen A dan 5 genogrup untuk segmen B.
- Khususnya, genogrup A2 terdiri dari 4 garis keturunan yang berbeda.
- Dalam klasifikasi baru ini, genotipe A1B1, A2B1, A3B2, dan A8B1 masingmasing sesuai dengan fenotipe klasik, varian, sangat virulen, dan yang dilemahkan.
PATOGENESIS DAN PRESENTASI KLINIS
Dalam patogenesis khasnya, IBDV memasuki tubuh melalui jalur pernapasan atau fekal-oral, tempat awalnya ia bereplikasi dalam makrofag dan sel limfoid di usus atau area sekitarnya.
- Replikasi awal ini memicu viremia primer melalui sirkulasi portal, yang memungkinkan virus mencapai organ target utamanya, bursa Fabricius.
- Di dalam bursa, virus secara aktif bereplikasi dalam folikel dan limfosit B, yang secara aktif membelah pada anak ayam.
- Infeksi ini menyebabkan degenerasi dan nekrosis folikel, terutama menyerang limfosit B IgM+, dan menyebabkan infiltrasi heterofil.
- Heterofil ini akhirnya mengalami nekrosis dan difagositosis.
Di daerah interfolikel, hiperplasia sel retikuloendotelial diamati, mengakibatkan atrofi bursa yang progresif (Müller et al., 2012).
Meskipun virus tidak bereplikasi dalam limfosit T, apoptosis sel-sel ini diamati di timus, dengan pemulihan lesi mikroskopis terjadi beberapa hari setelah infeksi. (Jagdev dan kawan-kawan, 2000).
- Mengenai viremia sekunder, hal itu dimulai sekitar 11 jam setelah replikasi di bursa.
Selama fase ini, virus memasuki aliran darah dan menyebar ke ginjal, otot, dan organ lainnya, yang mengakibatkan tandatanda klinis seperti depresi, bulu kusut, anoreksia, dan diare.
Dalam kasus yang parah, hal ini dapat mengakibatkan kematian. ( (Eterradossi dan Saif, 2008).
Virus ini menstimulasi limfosit B, meningkatkan ekspresi gen antivirus dalam jalur interferon tipe I (IFN), gen proapoptotik, dan sitokin proinflamasi.
- Selain itu, protein VP2 dan VP5 menginduksi apoptosis pada limfosit B dan sel limfoid lainnya.
Selama replikasi virus, terjadi infiltrasi limfosit T yang signifikan ke dalam bursa, yang bertahan hingga sekitar 12 minggu pasca infeksi.
- Sejak hari ke-7 pasca infeksi, limfosit T CD8+ (sitotoksik) jumlahnya melebihi limfosit T CD4+ secara proporsional dengan limfosit T CD4+.
- Peningkatan limfosit T CD8+ ini mendorong pembunuhan sel dengan melisiskan sel yang mengekspresikan antigen virus dan dengan memproduksi sitokin proinflamasi, seperti IFN-γ.
- Sitokin ini memicu pelepasan oksida nitrat oleh makrofag, meningkatkan kerusakan jaringan bursa (Jagdev et al., 2000) dan berkontribusi terhadap kelelahan sel imun.
Varian virus menginduksi peningkatan kadar IFN-γ, IL-6, IL-8, IL-18, NLRP3, caspase 1, dan TNF-α, yang meningkatkan peradangan dan mengubah lingkungan mikro jaringan.
- Strategi ini menekan aktivitas limfosit B, sehingga memungkinkan virus untuk menghindari respons imun, sehingga mengakibatkan peningkatan kerusakan bursa dan imunosupresi yang lebih parah dibandingkan dengan strain klasik. ( (Jagdev dkk., 2000; Li dkk., 2023).
Misalnya, Li et al. (2023) menunjukkan bahwa vvIBDV menunjukkan patogenisitas yang tinggi, efisiensi replikasi yang ditingkatkan, dan kapasitas yang signifikan untuk merusak bursa dan jaringan lain, yang menyebabkan tingkat kematian yang tinggi.
- Strain ini menyebar ke bursa, tonsil sekum, timus, dan limpa, dan mengubah kadar sitokin dalam bursa.
- Sebaliknya, varian SHG 19 dari Cina, yang dilaporkan pada tahun 2020, menunjukkan replikasi yang berkurang dan tidak menyebabkan kematian tetapi tetap menyebabkan kerusakan organ imun yang parah mirip dengan vvIBDV.
Varian ini menyebabkan nekrosis luas dan disintegrasi limfosit B, meskipun perubahan ini berkembang lebih lambat—sekitar 12 jam lebih lambat dibandingkan dengan strain yang sangat virulen.
- Mekanisme yang tertunda ini dapat berkontribusi terhadap kemampuan varian untuk mengeluarkan virus secara eksternal, sehingga berpotensi menjadikannya strain epidemik yang dominan (Fan et al., 2020).

TANTANGAN UNTUK VAKSIN KOMERSIAL
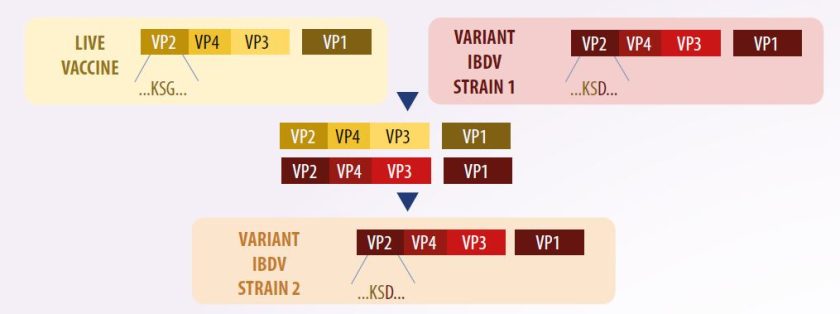
Vaksinasi dengan genotipe yang berbeda dari virus tipe wild dapat menyebabkan keragaman genetik di antara strain virus yang beredar.
- Dalam kasus seperti itu, reassortment dapat terjadi – seperti antara segmen A yang sangat virulen dan segmen B dari strain klasik – yang mengakibatkan kematian hingga 80% pada ayam dengan lesi bursal akut (Pikuła et al., 2018) (Gambar 3).
Selain itu, jarak antigenik antara galur virus tipe wild dan galur vaksin berarti bahwa varian-varian tersebut mungkin tidak dapat dikontrol secara efektif oleh vaksin serotipe 1 konvensional.
- Oleh karena itu, sekarang direkomendasikan agar produksi vaksin untuk melawan IBDV menyertakan pemetaan antigenik, sebuah metode komputasi yang digunakan untuk menentukan jarak antigenik antar strain. Teknik ini telah berhasil diterapkan pada virus influenza kuda dan manusia.
- Di sisi lain, penting untuk menilai perlindungan silang selama proses pengembangan vaksin untuk memastikan kemanjurannya (Boudaoud et al., 2016).
KESIMPULAN
- IBD masih menjadi tantangan yang signifikan bagi industri unggas karena mutasi virus yang cepat, perubahan genetik, dan munculnya varian baru yang sangat mematikan.
- Varian-varian ini berkontribusi pada penekanan imun yang parah, tingkat kematian yang tinggi, dan peningkatan kerentanan terhadap infeksi sekunder, meskipun ada upaya vaksinasi.
- Kompleksitas genetik virus, terutama pada protein VP2, mempersulit pengembangan vaksin yang efektif, karena variabilitas antigenik dapat merusak kemanjuran vaksin.
- Evolusi virus yang sedang berlangsung membutuhkan strategi yang lebih canggih, seperti pemetaan antigenik dan penilaian perlindungan silang, untuk meningkatkan desain vaksin dan meningkatkan langkahlangkah pengendalian terhadap penyakit yang berdampak besar secara ekonomi ini.
*Referensi berdasarkan permintaan kepada penulis

