



En el currículo médico, la patología general es la transición entre las disciplinas básicas y las clínicas. Los métodos aplicados al estudio de la anatomía, la histología y la fisiología, se utilizan para detectar la biología alterada y sus causas.

En medicina veterinaria, la patología general tiene alcances más amplios porque además de la transición descrita, representa una introducción a la patología comparada ya que se estudian varias clases de vertebrados e incluso invertebrados como el caso de las enfermedades de las abejas y los camarones que forman parte de la producción de alimentos para el consumo humano y por lo tanto caen dentro del campo de estudio de las ciencias veterinarias.
La patología aviar tiene una categoría diferente y específica porque el patólogo aviar es un profesional de muy alta especialidad que combina en la misma persona al clínico y al zootecnista por las características de la industria avícola.
Durante la segunda mitad del Siglo XX, la avicultura pasó de ser una actividad complementaria en la economía rural, generalmente en forma de pequeñas y medianas empresas familiares, a una industria de alta tecnología y fuertes inversiones que alcanzó una participación prioritaria en el mercado mundial de alimentos.
-Al iniciar el siglo XXI atestiguamos como los programas de control de las enfermedades infecciosas de las aves alcanzaron su dimensión global y el auge en el comercio internacional de aves y sus productos generó la presentación de los mismos problemas de sanidad animal y salud pública en todas las zonas avícolas del mundo.
-Este reto formidable ha sido enfrentado por los patólogos aviares a través de la adopción de los permanentes avances en la detección y caracterización de los agentes patógenos que infectan a las aves.
- Mediante el perfeccionamiento de los métodos de cultivo, aislamiento, caracterización y detección inmunológica y también con el uso extensivo de los procedimientos basados en la genética molecular.
Se han difundido ampliamente los buenos resultados en la prevención y control de enfermedades en condiciones de alta concentración de animales y producción masiva que caracterizan a la industria avícola moderna.
- Debido a que los avances logrados mejoraron la especificidad y sensibilidad de los métodos de diagnóstico y acortaron los tiempos de prueba para obtener resultados rápidos y confiables.
Existe, sin embargo, un aspecto mucho menos difundido, pero igualmente relevante que implica que este esfuerzo creó paralelamente modelos de alto valor para el avance del conocimiento de las ciencias biomédicas.
El propósito de este artículo es resaltar algunos ejemplos relevantes de las múltiples aportaciones de la patología aviar al avance de la medicina.
LA BURSECTOMÍA DE GLICK
En 1956, Bruce Glick y Timothy Chang demostraron que las respuestas de producción de anticuerpos se suprimen en las aves bursectomizadas y este estudio se convirtió en la piedra angular de la inmunología moderna.
- El significado de la Bolsa de Fabricio en la transformación de los linfocitos B con la consecuente población tisular de células plasmáticas y producción de anticuerpos se demostró con los experimentos de bursectomía al nacimiento realizados por Glick.

Los pollos con bursectomía seguida de irradiación al día de edad, no tienen células plasmáticas en los tejidos ni forman anticuerpos, pero su inmunidad celular permanece funcional mientras el timo se conserve intacto.
La mayor actividad de la bolsa ocurre el primer día de edad cuando es poblada por células troncales provenientes del hígado, del bazo y de la médula ósea y estas células reciben en la bolsa la inducción genética que los transforma en linfocitos B.
Actualmente ha quedado demostrado que en dicha inducción genética:
- Se activan las secuencias de síntesis de IgM que queda ligada a la membrana citoplásmica; y
- Un marcador distintivo de los linfocitos B y su sitio de reconocimiento antigénico.
Después de varias generaciones de replicación celular en la Bolsa de Fabricio, los linfocitos B migran a otros tejidos para formar los agregados linfoides foliculares.
- Estos linfocitos, al ser estimulados antigénicamente sufren transformación blástica, proliferación clonal y transformación en células plasmáticas productoras de las inmunoglobulinas.
En los mamíferos, las Placas de Peyer del intestino se consideran el equivalente de la Bolsa de Fabricio de las aves.
- Son islas de linfocitos localizadas en la submucosa del íleon terminal cuya remoción completa es imposible quirúrgicamente aún con el uso adicional de radiaciones y de ahí la enorme relevancia del modelo aviar.
Los trabajos de Glick y Chang desencadenaron los estudios de identificación de los órganos linfoides de diferenciación linfocitaria que generaron la noción que existe:
- Un sistema de inmunidad humoral y
- Otro de inmunidad celular basados respectivamente en linfocitos B y linfocitos T.
Siendo los modelos aviares los precursores porque las aves permiten una separación nítida de los órganos linfoides centrales de diferenciación.
En 1975, Le Douarin y Jotereau, realizaron un seguimiento de las células linfoides del timo en la vida embrionaria del pollo en quimeras interespecíficas.
En el embrión de pollo, el timo es funcional a partir del séptimo día de incubación.
- La población celular del timo de 6 días es básicamente epitelial.
- Mientras que a partir del día 7 las células predominantes son linfoblastos.
La timectomía al nacimiento en el pollo no inhibe la población de células plasmáticas en los tejidos ni la producción de anticuerpos, pero si compromete seriamente las respuestas de inmunidad celular.
Es así como los modelos aviares demostraron definitivamente que:
- El timo es responsable de la diferenciación de los linfocitos T, protagonistas de la inmunidad celular y
- La Bolsa de Fabricio es el órgano de diferenciación de los linfocitos B que dan origen a las células plasmáticas productoras de inmunoglobulinas.
CONTROL GENÉTICO CELULAR
El 25 de abril de 1953 se publicó uno de los descubrimientos más relevantes de las ciencias biológicas intitulado “Estructura molecular de los ácidos nucléicos. Una estructura para el ácido desoxirribonucleico”, por James Watson y Francis Crick.
Desde entonces se sabe que los ácidos nucéicos constituyen el material genético de todos los organismos conocidos y se inició la era de la biología molecular.
Sin embargo, las técnicas de biología molecular se fueron desarrollando en un inicio con mucho menos celeridad y tomó 8 años el establecimiento de un modelo de control genético celular que fue el modelo del operador desarrollado por Jacob y Monod del Instituto Pasteur en 1961.
Este se basa en el descubrimiento de que en las bacterias, las enzimas específicamente necesarias para la utilización de un nutriente:
- Se producen por los organismos bacterianos sólo en aquellos casos en donde los medios de cultivo contienen dicho nutriente.
- Este fue el estudio piloto de los caminos del control genético celular y se basa en el gen lac de Escherichia coli.

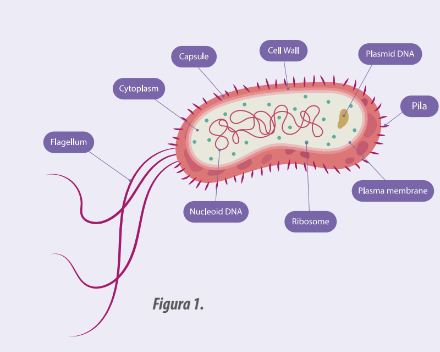
Figura 1: Diagrama de la estructura de las bacterias de Escherichia Coli.
El gen lac es el que se expresa con la síntesis de la enzima β galactosidasa que desdobla la lactosa en glucosa y galactosa.
Aquí entra en juego el modelo aviar porque E. coli tiene una proteína que se denominó represor que se encuentra bloqueando al operador que es el sitio donde se activa la ARN polimerasa en el genoma bacteriano para iniciar la transcripción del ARNm que corresponde a la secuencia del gen lac.
- El inductor del sistema es directamente la lactosa que tiene una afinidad química con el represor que lo liga liberando así al operador e iniciándose la transcripción.
- El ARNm transcrito pasa al citoplasma y se traduce en los ribosomas para la traslación y encadenamiento de los aminoácidos que forman los péptidos de la β galactosidasa.
- Al desdoblar la lactosa en glucosa y galactosa, se libera al represor que vuelve a su posición de bloqueo del operador.
Este elegante modelo de control genético celular generó múltiples ensayos para encontrar modelos equivalentes reproducibles experimentalmente en animales superiores.

