



Conteúdo disponível em:


A busca contínua por melhoria na alimentação de aves de produção tem levado a inclusão de ervas e especiarias na dieta, com o objetivo de substituir aditivos alimentares, reduzir a utilização de antimicrobianos, ou até mesmo para a utilização de alimentos alternativos, onde seus componentes não possuem pigmentantes na composição.
Exemplos são os extratos de plantas, óleos essenciais e aditivos fitogênicos. Estes produtos apresentam compostos biologicamente ativos como polifenóis, carotenoides e vitaminas, que podem atuar como antioxidantes, pigmentantes ou imunomoduladores, apresentando esses efeitos na ração, nas aves, nos ovos ou até mesmo na progênie.


Antioxidantes ajudam a prevenir o estresse oxidativo através da remoção de intermediários de radicais livres e são, portanto, um agente efetivo contra várias doenças. Em nutrição animal, as propriedades antioxidantes são apreciadas por agir principalmente como promotores de saúde (Faehnrich et al., 2016).
Os carotenoides
Os carotenoides são substâncias com características pigmentantes encontradas em organismos fotossintetizantes e não fotossintetizantes, plantas superiores, algas, fungos, bactérias e em alguns animais. Essas substâncias podem ser encontradas com cores entre o amarelo e o vermelho, com mais de 600 estruturas já caracterizadas (Fraser, 2004).
Nutricionalmente os carotenoides são classificados em:
- carotenoides pró-vitamínicos, representando os que têm atividade de pró-vitamina A, ou;
- carotenoides inativos, que desempenham apenas atividade antioxidante ou corante (Olson, 1999).
Já quimicamente, podem ser divididos em dois grupos:
- carotenoides hidrocarbonados, denominados carotenos; e
- carotenoides oxigenados, denominados xantofilas (Goodwin, 1965).
Estes dois grupos principais podem ser estruturalmente subdivididos em sete grupos, com seus principais representantes:
- Hidrocarbonetos – carotenos (α, β, γ e δ) e pelo licopeno;
- Álcoois – xantofilas (criptoxantina, zeaxantina e luteína);
- Cetonas – equinenona, cantaxantina e astacina;
- Epóxidos – flavoxantina;
- Éteres – espiriloxantina;
- Ácidos – crocetina;
- Ésteres – bixina e crocina;


Como os carotenoides são substâncias lipossolúveis, o processo de absorção e mecanismos de transporte celular são similares ao dos lipídeos. Assim, os carotenoides atravessam a membrana celular via lipoproteínas e se acumulam em células de diversos tecidos. Nas poedeiras em particular são transferidos em grande quantidade para o conteúdo da gema do ovo (Faehnrich et al., 2016).
Desta forma, à medida em que o teor de carotenoide na ração aumenta, a sua concentração na gema do ovo aumenta na mesma proporção.

Nos processos de oxidação que ocorrem nos ovos, os lipídios presentes na gema são oxidados em radicais livres (Ren et al., 2013). A reação de radicais livres com ácidos graxos poli-insaturados inicia um processo em cadeia conhecido como peroxidação lipídica. Os ácidos graxos poli-insaturados presentes na gema dos ovos com cadeias longas são particularmente suscetíveis à peroxidação (Pappas et al., 2005; Mohiti-Asli et al., 2008), o que altera a estrutura das membranas celulares, provocando modificações na estrutura dos aminoácidos, mudanças nas atividades enzimáticas e atingindo o DNA (Lima e Abdalla, 2001).
A geração de radicais livres no organismo constitui uma ação fisiológica contínua. Quando a produção supera a remoção, estes se acumulam causando o chamado “estresse oxidativo” que pode lesar as células, se esse acúmulo for mantido no organismo por período prolongado. Para se defender dos radicais livres, o organismo utiliza os antioxidantes que podem ser endógenos, como a enzima superóxido dismutase (SOD) por exemplo, ou exógenos, como os carotenoides obtidos na alimentação (Machado et al., 2006; Lima et al., 2012). Os carotenoides acíclicos, que possuem nove ou mais duplas ligações conjugadas, são capazes de retirar do meio espécies altamente reativas de oxigênio (Mcbride, 1996).
Desta forma, a ingestão de carotenoides pelas aves pode aumentar a capacidade antioxidante total no organismo e sua transferência para o conteúdo da gema leva a proteção do conteúdo lipídico da mesma, protegendo o ovo dos efeitos prejudiciais das espécies reativas de oxigênio e dos radicais livres (Sies; Stahl, 2003).
Hoje vários produtos à base de carotenoides têm sido utilizados na nutrição animal com esse objetivo. A capacidade de sequestrar o oxigênio por parte dos carotenos e xantofilas em ordem crescente é: licopeno, astaxantina ou cantaxantina, β-caroteno ou bixina, luteína e crocina (Fontana et al., 2000).
Xantofilas em produção de aves

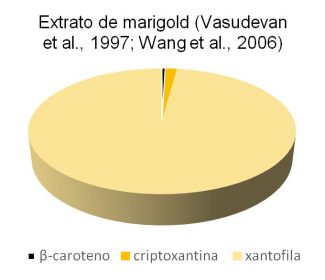
Os principais pigmentos presentes no extrato de marigold (Tagetes erecta L.) são os flavonóides e os carotenoides. O extrato de marigold contém aproximadamente 27% de carotenoides (Vasudevan et al., 1997; Wang et al., 2006), dos quais:
- 0,4% de β-caroteno,
- 1,5% de éster de criptoxantina e
- 86,1% de éster de xantofila

O extrato de marigold contém 12 g/kg de xantofilas, dos quais 80 a 90% são de luteína, sendo um carotenoide amarelo (Galobart et al., 2004) e normalmente, a luteína representa aproximadamente metade do total de xantofilas na gema do ovo (Steinberg et al., 2000).

Em função das características apresentadas para o extrato de marigold (Tagetes erecta L.), realizou-se um trabalho de pesquisa para identificar os efeitos da suplementação de xantofila (luteína), na dieta de matrizes de codornas.
A inclusão em níveis crescentes do extrato de marigold na dieta não influenciou as variáveis de desempenho e qualidade de ovos estudadas (P>0,05). Já a luminosidade e teor de vermelho apresentaram efeito linear (P<0,05) crescente em função da inclusão dos níveis de extrato de marigold, evidenciando que os carotenoides foram transferidos para a gema (Staub et al., 2018). Desta forma, acredita-se que sua maior concentração nos ovos férteis pode aumentar a capacidade antioxidante nos ovos e produzir pintinhos de melhor qualidade.
Surai e Sparks (2001) compararam ovos, embriões e pintos produzidos por matrizes alimentadas com dieta à base de milho, rica em carotenoides, especialmente luteína e zeaxantina (11,8mg de carotenoides/kg) e à base de trigo (5,6mg de carotenoides/kg). Os ovos provenientes de galinhas alimentadas com dieta à base de milho apresentaram maiores (P≤0,01) concentrações de beta + gama-tocoferol, carotenoides totais, luteína e zeaxantina, assim como os tecidos dos pintos nascidos destes ovos.
Os autores concluíram que a dieta da matriz tem importante função na formação do sistema antioxidante durante o desenvolvimento embrionário e que a dieta à base de milho aumenta o potencial antioxidante da gema do ovo e dos tecidos embrionários, quando comparada à dieta à base de trigo.
Considerações finais
Dietas ricas em substâncias antioxidantes, principalmente carotenoides, possuem o potencial de melhorar o status oxidativo do metabolismo geral das aves e de seus ovos. Esses efeitos se refletem na condição de saúde e bem-estar das matrizes e da progênie, contribuído para a produção de ovos férteis de qualidade e influenciando o desenvolvimento embrionário e o desempenho do pintinho pós-eclosão.
Esse conteúdo foi útil para você? Cadastre-se para receber nossa newsletter semanal!
Referências
Faehnrich, B., Lukas, B., Humer, E., Zebeli, Q. Phytogenic pigments in animal nutrition: potentials and risks. Revisão, J Sci Food Agric; 96: 1420–1430, 2016.
Fontana, J.D.; Mendes, S. V.; Persike, D.S.; Peracetta, L.; Passos, M. Carotenóides Cores Atraentes e Ação Biológica. Biotecnologia Ciência e Desenvolvimento, nº13, p.40-45, março/abril, 2000.
Fraser PD, Bramley P. M.The biosynthesis and nutritional uses of carotenoids. Prog Lipid Res 43:228–265, 2004.
Galobart, J., Sala, R., Rincón-Carruyo, X., Manzanilla, E.G., Vil, B., Gasa, J. Egg yolk color as affected by saponification of different natural pigmenting sources. J. Appl. Poultry Res. 13, 328–334, 2004.
Goodwin, T. W.. Chemistry and biochemistry of plant pigments. Academic Press. 1965.
Griffin, H. D.; Perry, M. M.; Gilbert, A. B. Yolk formation. Physiology and Biochemistry of the Domestic Fowl. London, UK: Academic Press, v. 5, p. 345-380, 1984.
Lima, E.S. & Abdalla, D.S.P. Peroxidação lipídica: mecanismos e avaliação em amostras biológicas. Brazilian Journal of Pharmaceutical Sciences. Vol. 37, n. 3, 2001.
Lima, J. P.; Lopes, C. O.; Dias, N. A. A.; Pereira, M. C. A. Atividade e biodisponibilidade dos carotenóides no organismo (artigo de revisão). Revista Ciências em Saúde, Lavras v. 2, n.1, p. 65-73, 2012.
Mcbride, J. It plants pigments paint an antioxidants substance rainbow. Agricultural Research, Washington, vol.44, Iss. 11, p.4-8, nov, 1996.
Mohiti-Asli M, Shariatmadari F., Lotfollahian H. The influence of dietary vitamin E and selenium on egg production parameters, serum and yolk cholesterol and antibody response of laying hen exposed to high environmental temperature. Arch Geflugelk. 74: 43-50, 2010.
Nachtigall, A. M. Extração, saponificação e atividade antioxidante de luteína obtida de flores de Tagetes pátula L. e Calendula officinalis L. Tese (Doutorado em Ciência e Tecnologia de Alimentos) – Faculdade de Engenharia de Alimentos, Universidade Federal de Viçosa, Viçosa, 2007.
Oliveira, D.D; Baião, N.C; Cançado, S.V. Fontes de lipídeos na dieta de poedeiras: Desempenho produtivo e qualidade dos ovos. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.62, n. 3, p. 718 – 724. 2010.
OliveirA, N. T. E.; Fonseca, J. B.; Soares, R. T. R. N.; Ferreira, K. S.; ThiebauT, T. L. Pigmentação de gemas de ovos de codornas japonesas alimentadas com rações contendo colorífico. Ciência e Agrotecnologia, v.31, n.5, p.1525-1531, 2008.
Olson, J. A. Absorption, transport, and metabolism of carotenoids in humans. Pure & Applied Chemistry (IUPAC), Great Britain, vol.66, nº5, p.1011-1016, 1994.
Pappas, A. C.; Acamovic, T.; Sparks, N. H. C.; Surai, P. F.; Mcdevitt, R. M. Effects of supplementing broiler breeder diets with organic selenium and polyunsaturated fatty acids on egg quality during storage. Poultry Science, v. 84, p. 865–874, 2005.
Quackenbush FW, Miller SL. Composition and analysis of the carotenoids in marigold petals. J Assoc Off Anal Chem, 55:617–21, 1972.
Ren Y., Perez T.I., Zuidhof M.J., Renema R.A., Wu J. Oxidative stability of omega-3 polyunsaturated fatty acids enriched eggs. Journal of Agricultural and Food Chemistry, 61: 11595–11602, 2013.
Ribeiro, E. P. & Seravalli, E. A. G.. Química de Alimentos. Instituto Mauá de Tecnologia. Editora Edgard Blücher Ltda, 1ª edição, São Paulo, p.155-157, 2004.
Rivas J. D. L. Reversed-phase high-performance liquid chromatographic separation of lutein and lutein fatty acid esters from marigold flower petal powder. J Chromatogr A, 464:442–7, 1991.
Sies, H., Stahl, W. Vitamins E and C, b-carotene, and other carotenoids as antioxidants. American Journal of Clinical Nutrition, Bethesda, v.62, n.6, p.1315-1321, 1995.
Staub, L., Roza L. D. F., Menezes de Oliveira, E., Sesco de Mendonça, M., Selan, N., dos Santos T. C.;. Performance and quality of eggs of Japanese quail breeders supplemented with xanthophylls. 2nd Latin American Scientific Conference, 2018, Campinas, SP. Poult. Sci. 97(E-Suppl. 2). Disponível em: https://www.poultryscience.org/latin18/abstracts/77.pdf Acesso: 30/01/2019.
Surai, P. F.; Sparks, N. H. C. Designer eggs: from improvement of egg composition to functional food. Trends in Food Science & Technology, London, v. 12, p. 7-16, 2001.
Vasudevan, P.; Kashyap, S.; Sharma, S. Tagetes: a multipurpose plant Bioresour. Technol., v.62, p.29-35, 1997.
Wang, M.; Tsao, R.; Zhang, S.; Dong, Z.; Yang, R.; Gong, J.; Pei, Y. Antioxidant activity, mutagenicity/anti-mutagenicity, and clastogenicity/anti-clastogenicity of lutein from marigold flowers. Food and Chemical Toxicology, v. 44, p. 1522-1529, 2006.