



Nội dung có ở: English Indonesia (Indonesian) Melayu (Malay) ไทย (Thai) Philipino
Vi-rút Avian Influenza disease Virus (AIV) thuộc loài virus cúm type A, họ Orthomyxoviridae, chi Alphainfluenzavirus (theo phân loại của Ủy ban Phân loại Vi-rút Quốc tế – International Committee of Viral Taxonom).
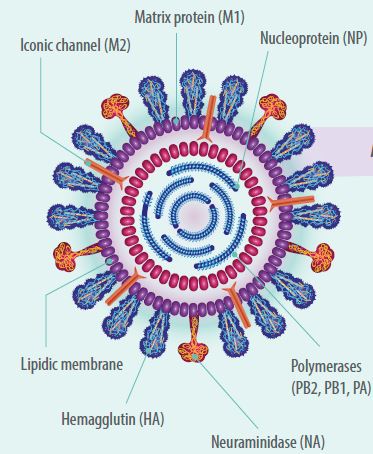

- AIV có đặc trưng là được bao quanh bởi một lớp màng phospholipid và có dạng hình cầu hoặc hình sợi với kích thước khoảng 80-120nm.
- Vi-rút có một bộ gen bao gồm 8 phân đoạn RNA mạch đơn thẳng (linear single-stranded RNA) có chiều 3-5′ (âm).
- Bộ gen RNA mã hóa cho 11 protein, chín trong số đó là protein cấu trúc/structural proteins (PB2, PB1, PB1-F2, PA, HA, NA, M1 và M2) và hai protein phi-cấu trúc/non-structural proteins (NS1 và NS2).
Các phân đoạn là:- Phân đoạn Một mã hóa cho enzyme polymerase PB2;
- Phân đoạn Hai mã hóa cho enzyme polymerase PB1 hoặc PB1-F2;
- Phân đoạn Ba mã hóa cho enzyme polymerase axit PA.
- Phân đoạn Bốn mã hóa cho glycoprotein kết dính (adhesion glycoprotei) gọi là hemoagglutinin (HA) – liên quan đến sự liên kết vi-rút với tế bào, xác định độ độc lực và là kháng nguyên cung cấp sự phân loại vi-rút influenza A thành 18 loại HA khác nhau (16 loại HA ở chim và 2 loại HA ở dơi).
- Phân đoạn Năm mã hóa cho nucleoprotein (N) và là kháng nguyên cung cấp sự phân loại vi-rút AIV theo chi thành A, B, C và D.
- Phân đoạn Sáu mã hóa cho neuroaminidase (NA) – một glycoprotein có trên bề mặt của vi-rút liên quan đến việc giải phóng các hạt vi-rút khỏi thụ thể của tế bào vật chủ và là kháng nguyên cung cấp sự phân loại vi-rút influenza type A thành 11 NA riêng biệt (9 NA ở chim và 2 NA ở dơi).
- Phân đoạn Bảy cho ma trận/matrix (M1 và M2).
- Cuối cùng, phân đoạn Tám mã hóa cho protein phi-cấu trúc NS1 và NS2.
- Các vi-rút AIV có ở các loài chim thì có thể biểu hiện trên bề mặt của chúng một trong 16 HA và một trong 9 NA, mà trên lý thuyết có thể tạo ra 144 phân nhóm (subtype) virus. Hai loại protein này có các biến thể kháng nguyên thông qua hai cơ chế sau:
- Cơ chế đầu tiên là sự chuyển dịch kháng nguyên (antigenic drift) bao gồm các đột biến ba-zơ (base mutation) (thay thế, chèn, xóa hoặc đảo ngược) do thiếu sự điều chỉnh của enzyme RNA polymerase trong quá trình tổng hợp bộ gen của virus.
- Cơ chế thứ hai là bằng cách tái tổ hợp các gen phân đoạn của vi-rút khi một tế bào bị nhiễm bởi hai phân nhóm khác nhau. Cơ chế thứ hai cho giúp vi-rút có được các phân đoạn bộ gen từ các loài khác như heo và người.
VẬT CHỦ TỰ NHIÊN CỦA AIV
Hiện nay có 16 HA và 9 NA được tìm thấy trong tự nhiên cùng tồn tại với khoảng 225 loài thủy cầm hoang dã phân bố trên khắp thế giới và chủ yếu thuộc bộ Anseriforme (vịt, ngỗng và thiên nga) và bộ Charadriiforme (mòng biển, chim nhạn biển/terns và chim bờ biển/shorebirds).
- Những loại vi-rút này cũng được tìm thấy ở vùng đất ngập nước (wetlands), là môi trường sống dưới nước và nơi nước được xem là phương tiện lây truyền vi-rút, nhưng chỉ có một số ít mẫu phân lập vi-rút từ trong nước cho thấy khả năng truyền lây qua đường này rất hạn chế.
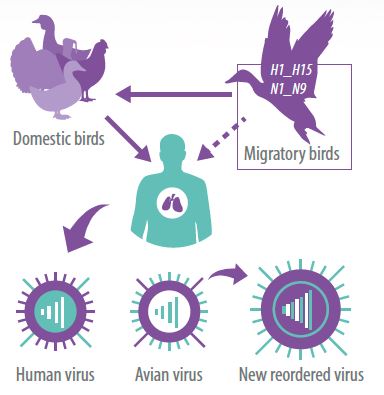

Ở Bắc Mỹ, Đức và Thụy Sĩ, các phân nhóm có tần suất cao nhất ở vịt là H4N6 và H6N2, trong khi các phân nhóm H13-H16 đã được báo cáo phát hiện ở các loài chim bờ biển.
Tuy nhiên, một số phân nhóm liên quan đến hemoagglutinin H5, H6, H7 và H9 có nguồn gốc từ các loài thủy cầm hoang dã là những loại vi-rút độc lực thấp và thường xuyên tiếp xúc với hệ thống trang trại gia cầm như vịt, gà và gà tây; các vi-rút này bắt đầu thích nghi với sự gia tăng độc lực và khả năng gây chết trên sinh vật chủ.
Một ví dụ về điều này là sự xuất hiện và hiện diện của phân nhóm H5N1 có độc lực cao trong gia cầm ở Trung Quốc, có thể gây chết gia cầm, một số loài chim hoang dã, động vật hữu nhũ và trong những trường hợp hiếm, gây chết cả con người khi tiếp xúc gần với gia cầm.
- Khi phát hiện vi-rút trong các trường hợp viêm phổi ở người tại Á châu, bộ gen có nguồn gốc từ chim, nhưng virus vẫn chưa tiến hóa để có thể lây truyền trực tiếp từ người sang người.
Một ví dụ khác về bệnh truyền nhiễm từ động vật là chủng H7N9 được phát hiện ở 14 trường hợp tử vong trong số 63 người ở Trung Quốc.
- Bộ gen của phân nhóm vi-rút này liên quan về mặt di truyền với các loại vi-rút gia cầm ở các chợ bán chim sống và chim bồ câu.
- Mặc dù các phân nhóm H5N1 và H7N9 có liên quan về mặt phát sinh loài (phylogenetically) với gia cầm, nhưng sự tiếp xúc gần là một yếu tố quan trọng, cũng như các yếu tố sức khỏe đặc thù khác của con người.
Tổ chức Y tế Thế giới (WHO) cũng xem xét H9N2 ở mức độ thấp hơn, loại virus có độc lực thấp ở gà nhưng đã được phát hiện ở người.
CƠ CHẾ SINH BỆNH CỦA VI-RÚT AVIAN INFLUENZA VIRUS H5N2 CÓ NGUỒN GỐC MEXICO
Cơ chế sinh bệnh (pathogenesis) của vi-rút AIV Mexico đầu tiên có mức độ gây bệnh thấp và cao đã được nghiên cứu trong các thí nghiệm in vivo (trong sinh vật sống). Trong các nghiên cứu này, người ta đã chứng minh rằng:
- Vi-rút AIV H5N2BP/1994 được chủng cho gà nuôi 4 tuần tuổi sẽ gây tỷ lệ chết thấp với các triệu chứng hô hấp ở mức độ trung bình.
- Vi-rút lây lan đến phổi, các cơ quan lympho và các cơ quan nội tạng, gây hoại tử mô và suy giảm tế bào lympho ở các cơ quan lympho;
- AIV H5N2AP cũng lây lan sang các cơ quan khác như tim, não, khí quản và hạch manh tràng (cecal tonsils) gây hoại tử, xuất huyết và mất lông mao ở biểu mô khí quản.
- Nguyên nhân chết của gà bị cấy nhiễm vi-rút AIV H5N2AP có liên quan đến tổn thương nội mạc mao mạch và giảm tiểu cầu dẫn đến tổn thương mạch máu lan rộng.
- Trong thí nghiệm, người ta cũng chứng minh được rằng vi-rút H5N2AP có thể tồn tại trong mạch máu ngoại vi 28 giờ sau khi cấy nhiễm và lên đến 72 giờ sau khi kết thúc thí nghiệm.
Nghiên cứu cơ chế gây bệnh
Nghiên cứu về cơ chế gây bệnh của vi-rút AIVH5N2 ở các loài gia cầm khác như vịt, chim choi choi (plover), gà tây, gà lôi và chim cút bị cấy nhiễm vi-rút qua đường tĩnh mạch không bị bệnh và không lây nhiễm cho những con gà không có mầm bệnh cụ thể cùng tồn tại với các loài chim hoang dã bị nhiễm bệnh.
- Trong một nghiên cứu khác trên cơ chế gây bệnh của AIV H5N2BP/2007 ở vịt nhà khi chúng được cấy nhiễm qua đường mũi và khí quản cho thấy loại vi-rút này không gây ra các triệu chứng lâm sàng ở vịt nhưng có thể được phân lập từ mẫu cuốn mũi (turbinates), thanh quản, khí quản và phổi trong 48 giờ đầu sau khi cấy nhiễm và gây tình trạng viêm với sự thâm nhiễm lympho bào từ nhẹ đến trung bình khi quan sát dưới kính hiển vi.
Nghiên cứu về động học bài tiết
Các nghiên cứu về động học bài tiết của dòng virus AIV H5N2BP/2007 của Mexico phát hiện bằng cách bài tiết virus ở hầu họng (oropharynx) và hậu môn (cloaca) ít nhất 21 ngày sau khi cấy nhiễm vào gà và vịt.
- Trong khi các tác giả khác đã báo cáo rằng việc cấy nhiễm loại vi-rút H5N2BP của dòng Mexico ở vịt được bài tiết từ hai đến mười bốn ngày sau khi cấy nhiễm qua đường hô hấp và từ 0 đến bảy ngày sau khi cấy nhiễm qua hậu môn.
Người ta cũng quan sát thấy rằng AIV H5N2BP/2007 có nguồn gốc từ Mexico cấy nhiễm ở vịt nhà có thể được bài tiết ban đầu qua đường tiêu hóa và sau đó qua đường hô hấp và ngược lại ở gà.
Đột biến của các nucleotide
Hầu hết các đột biến nucleotide được nghiên cứu ở vi-rút AIV đều tập trung ở vùng phân cắt gen HA (gene cleavage region) vì có thể xác định được sự khác biệt giữa trình tự a-xít amin HA từ một đợt dịch bởi vi-rút và từ vắc-xin bởi vi-rút, làm giảm khả năng bảo vệ.
Sự thay đổi đặc thù này về tính hướng đích (tropism) giữa mô hô hấp và mô tiêu hóa được quan sát thấy ở gà và vịt được cấy nhiễm H7N3BP, H7N2BP, H7N3BP và H7N9BP từ vịt trời, gà và gà tây nuôi. Để giải thích cho sự thay đổi về tính hướng đích, ta có thể liên tưởng đến:
- Sự đột biến nucleotide xảy ra ở gen HA và gen NA trong quá trình sao chép và lây truyền vi-rút ở động vật như đã được quan sát thấy ở phân nhóm H11N9 ở vịt trời.
- Hoặc vì nó có gen từ các phân nhóm vi-rút khác có sự tái tổ hợp giữa vi-rút gà và vịt; như trường hợp phân nhóm H9N2BP ở vịt nhà từ Hàn Quốc và phân nhóm H5N2BP ở ngỗng hoang dã ở Phi châu.


Sự tích tụ các a-xít amin ba-zơ
Sự tích tụ của các a-xít amin ba-zơ (basic amino acids) trong vùng phân cắt HA thì có liên quan đến độc lực cao của vi-rút ở gà cũng đã được phân tích, nhưng sự có mặt hoặc thiếu vùng a-xít amin ba-zơ này trong một số trường hợp tương ứng với vi-rút có độc lực thấp, cho thấy sự tham gia của các gen khác trong độ độc lực của vi-rút, như PB1-F2, PB2 và NS1, trong đó các đột biến đã được tìm thấy ở gốc puric hoặc pyrimidic liên quan đến độc lực.


TÌNH HÌNH CỦA VI-RÚT AIV Ở MEXICO
AIV H5N2
Vào ngày 23 tháng 5 năm 1994 khi lần đầu tiên phân lập được vi-rút AIV H5N2 độc lực thấp đã được báo cáo, và vào tháng 12 năm đó, lần đầu tiên phân lập được vi-rút AIV H5N2AP ở đàn gia cầm trong trang trại thương mại bởi Phòng thí nghiệm của Ủy ban Phòng chống Bệnh lở mồm long móng và các Bệnh động vật khác (Commission for the Prevention of Foot-and-Mouth Disease and Other Animal Diseases – CPA) Hoa Kỳ-Mexico. Vi-rút AIV H5N2AP đã được loại trừ vào tháng 6 năm 1995.
Tuy nhiên, cho đến nay, AIV H5N2BP vẫn còn phổ biến ở các trang trại thương mại và gà mái thả vườn, nhưng do khả năng gây bệnh thấp nên vi-rút không thể phát hiện được trên lâm sàng vì gia cầm có thể phục hồi.
Trong 24 năm hiện hữu của phân nhóm H5N2BP, không có báo cáo nào cho thấy vi-rút đã đột biến thành dạng gây bệnh cao và bộ gen của vi-rút vẫn có nguồn gốc từ gia cầm.
AIV H7N3
Vào tháng 6 năm 2012, phân nhóm AIV H7N3 với các đặc điểm sinh học và phân tử có khả năng gây bệnh cao đã xuất hiện lần đầu tiên tại các trang traị gia cầm sản xuất trứng thương phẩm tại Tiểu bang Jalisco. Sau khi được đã kiểm soát, phân nhóm vi-rút đã biến mất (về mặt dịch tễ) trong 17 tuần, xuất hiện lại vào tháng 1 năm 2013 và hiện hữu cho đến nay ở một số khu vực có trang trại chăn nuôi gia cầm quy mô hoặc chăn nuôi gia cầm tại nhà.
- Bộ gen hoàn chỉnh của phân nhóm AIV H7N3 có nguồn gốc phát sinh loài với AIV từ vịt trời ở Hoa Kỳ chứ không phải từ người hoặc heo.
Phân nhóm AIV độc lực cao H7N3 hiện diện ở gà mái tại Tiểu bang Jalisco đã được xác định về mặt kháng nguyên và phân tử trong kết mạc của hai công nhân trong trang trại gia cầm tại khu vực này, những người có biểu hiện viêm kết mạc mà không sốt hoặc bệnh đường hô hấp.
- Trong thực nghiệm, chủng vi-rút chưa thích nghi (unadapted) này gây chết ở chuột và dễ dàng lây truyền trực tiếp giữa các loài chồn, cũng như dễ dàng nhân lên trong các tế bào phế quản của con người.

Hiện tại, các phân nhóm H5 và H7 là các loại vi-rút cúm AIV phải được báo cáo cho SENASICA và OMSA. Đối với các loại vi-rút AIV được phân lập hoặc phát hiện ở các loài chim hoang dã tại Mexico, các phân nhóm vi-rút AIV type A có độc lực thấp H7N3, H6N2, H4N2 và H5NX đã được báo cáo ở các loài vịt trời di cư.


Hiện nay, vi-rút AIV H5N1 và H7N9 có nguồn gốc từ châu Á là các phân nhóm có ảnh hưởng lớn nhất đến sức khỏe con người và nguồn gốc của chúng là từ sự tiếp xúc gần giữa các loài gia cầm, vịt trời với những người chăn nuôi và buôn bán chúng.

