



El Virus de la enfermedad de Influenza Aviar (VIA) pertenece a la especie virus de influenza tipo A que pertenece a la familia Orthomyxoviridae y género Alfainfluenzavirus con base al Comité Internacional de Taxonomía Viral.

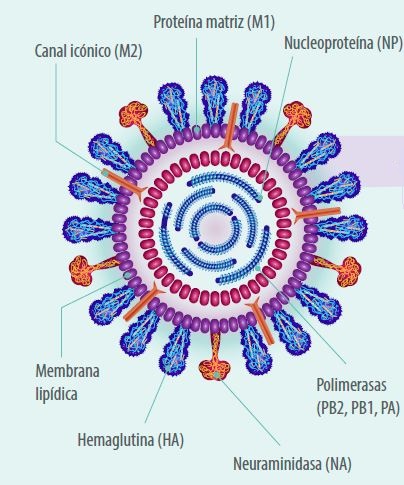
Figura 1. Estructura del virus de la Influenza Aviar.
-
- El VIA Se caracteriza por ser envuelto por una membrana de fosfolipidos y de forma esférica o filamentosa con un tamaño de aproximadamente de 80-120nm. ..
- Contiene un genoma compuesto por 8 segmentos de RNA lineal de cadena sencilla con sentido 3-5’ (negativo).
- El genoma de RNA codifica para 11 proteínas de las cuales nueve son estructurales (PB2, PB1, PB1-F2, PA, HA, NA, M1 y M2) y dos no estructurales (NS1 y NS2). Los segmentos son:
- El segmento uno codifica para la enzima polimerasa PB2;
- El segmento dos codifica para la enzima polimerasa PB1 o PB1-F2;
-
- El segmento tres codifica para la enzima polimerasa ácida PA.
- El cuarto segmento codifica para la glicoproteína de adhesión llamada hemoaglutinina (HA) que participa en la unión del virus a la célula, determina el grado de virulencia, y es el antígeno que permite clasificar a los virus de influenza tipo A en 18 HA distintas (16 HA en aves y 2HA en murciélagos).
- El quinto segmento codifica para la nucleoproteína (N) y es el antígeno que permite clasificar a los virus de influenza por género en A, B, C y D.
- El segmento seis codifica para la neuroaminidasa (NA) que es una glicoproteína presente en la superficie del virus que participa en la liberación de las partículas virales de los receptores de la célula del huésped y es el antígeno que permite clasificar al virus de influenza tipo A en 11 NA distintas (9 NA en aves y 2 NA en murciélagos).
- El segmento siete codifica para la matriz (M1 y M2).
- Por último, el segmento ocho codifica para la proteína no estructural NS1 y NS2.
-
- Los VIA que se encuentran en la clase aves pueden expresar en su superficie una de las 16 HA y una de las 9 NA que pueden originar en teoría 144 subtipos virales. Estas dos proteínas tienen variaciones antigénicas a través de dos mecanismos:
- El primero es la deriva antigénica que consiste en mutaciones por base (sustitución, inserción, eliminación o reversión) por la falta de corrección de la enzima RNA polimerasa durante la síntesis del genoma viral.
-
- El segundo es por medio de la recombinación de sus genes segmentados cuando una célula es infectada por dos subtipos diferentes. Este último mecanismo permite al virus adquirir segmentos genómicos de otras especies como cerdo y humano.

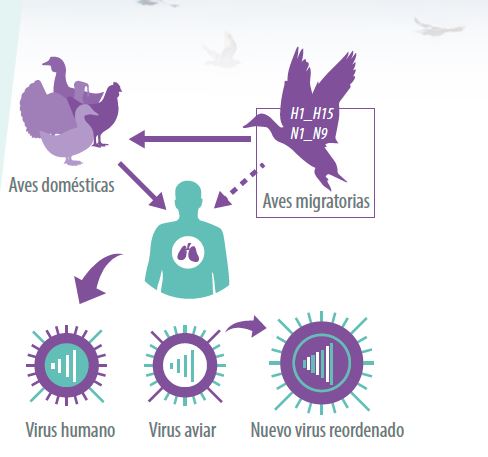
Figura 2. Detectar rápidamente la introducción de nuevos virus.
HUÉSPED NATURAL DEL VIA
Actualmente, las 16 HA y 9 NA se encuentran en la naturaleza en coexistencia con alrededor de 225 especies de aves acuáticas silvestres distribuidas en todo el mundo y que pertenecen principalmente al orden Anseriforme (patos, gansos y cisnes) y orden Charadriiforme (gaviotas, charranes y playeras).

Estos virus también se encuentran en los humedales que son el hábitat acuático y donde el agua es considerada un medio de transmisión, pero los pocos aislamientos en agua sugieren que es limitado.

En Norte América, Alemania y Suiza, los subtipos con mayor frecuencia en patos son el H4N6 y H6N2, mientras que subtipos del H13-H16 se han reportado en aves playeras.

Los virus presentes en aves silvestres son considerados de baja patogenicidad (BP) y tienen un limitado intercambio genético intercontinental.
Sin embargo, algunos subtipos relacionados a la hemoaglutinina H5, H6, H7 y H9 que provienen de aves acuáticas silvestres como virus de baja patogenicidad y que tienen contacto constante con sistemas de producción de aves domésticas de corral como son los patos, gallinas y pavos; comienzan un proceso de adaptación que se caracteriza por un incremento de la virulencia y capacidad para causar enfermedad fatal.
- Ejemplo de ello es el surgimiento y presencia actual del subtipo H5N1 de alta patogenicidad (AP) en aves de corral en China que ha mostrado su capacidad para matar aves domésticas, algunas aves silvestres, mamíferos y en raras ocasiones al humano cuando está en estrecho contacto con aves de corral.

Cuando el virus se ha detectado en casos de neumonía en humanos en Asia, el genoma tiene un origen aviar, pero el virus no ha evolucionado para transmitirse directamente de humano a humano.

Otro ejemplo de zoonosis ha sido el subtipo H7N9 que se detectó en 14 casos fatales de 63 humanos en China.
-
- El genoma de este subtipo está relacionado genéticamente con virus de aves de corral en mercados de aves vivas y palomas.
-
- Si bien el subtipo H5N1 y H7N9 han sido asociados filogenéticamente a las aves de corral, el estrecho contacto es un factor clave, así como otros factores de salud propios del humano.
La organización mundial de la salud también contempla en menor proporción al H9N2 que es de baja patogenicidad en pollos, pero que se ha detectado en humanos.
PATOGENIA DEL VIRUS DE INFLUENZA AVIAR H5N2 DE LINAJE MEXICANO
La patogenia de los primeros VIA mexicanos de baja patogenicidad y alta patogenicidad fueron realizados en experimentos in vivo. En estos estudios, ha evidenciado que:


- El VIA H5N2BP/1994 inoculado a pollos domésticos de 4 semanas de edad, causan baja mortalidad con moderados signos respiratorios.
- El virus se disemina al pulmón, órganos linfoides y órganos viscerales ocasionando necrosis de tejido y depleción de linfocitos en órganos linfoides;
- El VIA H5N2AP también se disemina a otros órganos como el corazón, cerebro, tráquea y tonsilas cecales en los que provoca necrosis, hemorragias y pérdida de cilios en epitelio de tráquea.
- La causa de muerte de los pollos infectados experimentalmente por el VIA H5N2AP se asocia al daño del endotelio capilar y trombocitopenia que generan un daño vascular generalizado.
- Experimentalmente también se mostró que el virus H5N2AP está presente en la sangre periférica a las 28 horas posteriores a la inoculación y hasta las 72 horas de terminado el experimento.
Estudio de patogenia


El estudio de patogenia del VIAH5N2 en otras especies de aves como el pato, chorlito, pavo, faisán y codorniz inoculados con el virus por vía endovenosa, no se enferman y no infectan a pollos libres de patógenos específicos que convivieron con las aves silvestres infectadas.
- En otro estudio la patogenia del VIA H5N2BP/2007 en patos domésticos infectados experimentalmente por vía intranasal e intratraqueal, mostró que este virus no ocasiona signos clínicos en el pato, pero se puede aislar de cornetes, laringe, tráquea y pulmones en las primeras 48 horas posteriores a la inoculación y ocasionar inflamación con infiltración linfocitaria leve o moderada.


Estudios de cinética de excreción
Los estudios de cinética de excreción del VIA H5N2BP/2007 de linaje mexicano se detectó la excreción del virus en orofaringe y cloaca al menos por 21 días posteriores a la inoculación en pollos y patos.
- Mientras que otros autores han reportado que la inoculación del subtipo H5N2BP de linaje mexicano en pato se excreta entre dos a catorce días posteriores a la inoculación por la vía respiratoria y de cero a siete días por la cloaca.
- También se observó que el VIA H5N2BP/2007 con linaje mexicano inoculado en pato doméstico puede tener una excreción inicialmente digestiva y posteriormente respiratoria y viceversa en pollo.
Esta característica de cambio de tropismo entre el tejido respiratorio y digestivo se ha observado en pollos y patos inoculados con H7N3BP, H7N2BP, H7N3BP y H7N9BP de patos silvestres, pollos y pavos domésticos. La explicación al cambio de tropismo se puede asociar a:
- Mutaciones de nucleótidos que ocurren en el gen HA y en el gen NA durante la replicación y transmisión viral en el animal como se ha observado en el subtipo H11N9 de pato silvestre.
- O porque tenga genes de otros subtipos de virus con los que haya existido recombinación entre virus de pollo y pato; como es el caso del subtipo H9N2BP en pato doméstico de Corea del Sur y el subtipo H5N2BP en gansos silvestres de África.
Mutaciones de nucleótidos
La mayoría de las mutaciones de nucleótidos estudiadas en el VIA se ha concentrado en la región de corte del gen HA porque se puede determinar la diferencia entre la secuencia de aminoácidos de la HA del virus de un brote y el virus de la vacuna, lo que disminuye la protección.

Acumulación de aminoácidos básicos
También se analiza la acumulación de aminoácidos básicos en la región de corte de la HA que se asocia a la alta virulencia del virus en los pollos, pero la presencia o falta de esta región de aminoácidos básicos en algunos casos corresponde a virus de baja patogenicidad; lo que sugiere la participación de otros genes en la virulencia como por ejemplo la PB1-F2, PB2 y NS1 donde se han encontrado mutaciones por base purica o pirimidica asociadas a la virulencia.

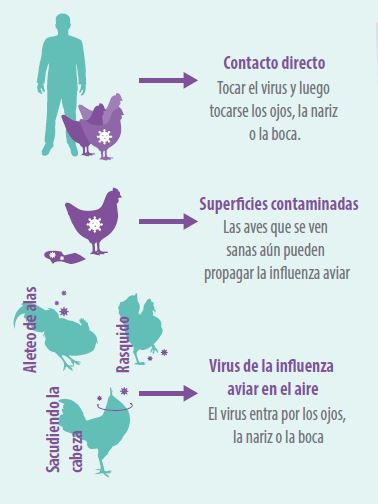
Figura 3. Infecciones humanas por virus de Influenza Aviar raras, pero posibles.


SITUACIÓN DE LOS VIRUS DE INFLUENZA AVIAR (VIA) EN MÉXICO
VIA H5N2
El 23 de mayo de 1994 se notificó el primer aislamiento del VIA H5N2 de baja patogenicidad (BP) y en diciembre de ese año el primer aislamiento del VIA H5N2AP en aves comerciales por el Laboratorio de la Comisión México-Estados Unidos para la Prevención de la Fiebre Aftosa y otras Enfermedades de los Animales (CPA). El VIA H5N2AP fue erradicado en junio de 1995.
Sin embargo, hasta la fecha el VIA H5N2BP prevalece en granjas comerciales y gallinas de traspatio, pero debido a que es de baja patogenicidad puede pasar desapercibido clínicamente ya que las aves se pueden recuperar.
En 24 años de presencia del subtipo H5N2BP, no hay reporte que demuestre que el virus ha mutado a la presentación de alta patogenicidad y su genoma es de origen aviar.
VIA H7N3
En junio de 2012 aparece por primera vez el VIA subtipo H7N3 con características moleculares y biológicas de alta patogenicidad en unidades de producción avícola productoras de huevo para consumo en el Estado de Jalisco. El cual después de ser controlado, estuvo ausente epidemiológicamente por 17 semanas, apareciendo en enero de 2013, y presente a la fecha en algunas regiones donde hay avicultura tecnificada o de traspatio.
- El genoma completo del VIA subtipo H7N3 tiene origen filogenético con VIA de patos silvestres de Estados Unidos y no con humanos o cerdos.

En el presente, el subtipo H5 y H7 son virus de influenza aviar que se deben notificar al SENASICA y OMSA. Con respecto a los VIA aislados o detectados en aves silvestres en México, se han reportado virus de influenza tipo A de baja patogenicidad del subtipo H7N3, H6N2, H4N2 y H5NX en patos silvestres migratorios.
El VIA subtipo H7N3 de alta patogenicidad presente en gallinas del Estado de Jalisco fue identificado antigénicamente y molecularmente en la conjuntiva de dos trabajadores avícolas en el Estado de Jalisco que presentaron conjuntivitis sin fiebre o enfermedad respiratoria.
- Experimentalmente, este aislamiento viral sin previa adaptación, causa enfermedad fatal en ratones y se transmite fácilmente de forma directa entre hurones, así como fácil replicación en células bronquiales humanas.


Figura 4. Virus de Influenza Aviar H5N1, impacto en la salud humana.
Actualmente el virus de influenza aviar H5N1 y H7N9 de linaje asiático son los subtipos con mayor impacto en la salud de las personas y su origen se dado por el estrecho contacto entre distintas especies de aves de producción, patos silvestres y las personas que los crían y comercian.

