
La extracción eficiente de energía dietética a partir de la grasa depende de una secuencia de eventos bien coordinada: emulsificación, hidrólisis enzimática, formación de micelas y absorción a través de la membrana del enterocito (1).
En condiciones comerciales, esta cascada fisiológica rara vez opera a máxima eficiencia.
El estrés térmico y la variabilidad en la calidad de las fuentes lipídicas reducen la digestibilidad de la grasa, generando una brecha energética que impacta directamente el índice de conversión alimenticia (ICA) y el rendimiento productivo.

LIPA® optimiza esta secuencia de eventos al suministrar un perfil concentrado y estandarizado de lisofosfolípidos (LFL), obtenidos mediante la hidrólisis enzimática de la lecitina de soja.
Para garantizar que cada inclusión aporte una dosis definida de lisofosfolípidos, el contenido activo de cada lote de producción se verifica por resonancia magnética nuclear (RMN).
La lecitina de soja, cuya fracción de fosfolípidos (PC) predominante es la fosfatidilcolina, ofrece una emulsificación moderada en su forma intacta.


La hidrólisis enzimática mediante la fosfolipasa A2 (PLA2) elimina una de las cadenas de ácidos grasos, transformándola eficientemente en lisofosfatidilcolina (LPC).
![]()
La relevancia comercial de esta modificación estructural es clara. El lisofosfolípido resultante cuenta con un balance hidrofílico-lipofílico (HLB) superior con respecto a los fosfolípidos. Su único ácido graso le confiere una geometría cónica, ideal para estabilizar micelas mixtas.
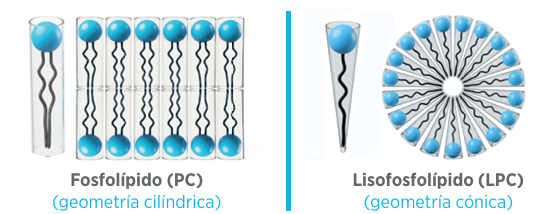
Estas propiedades fisico-químicas le permiten emulsificar eficazmente sin depender de la bilis endógena. Dicha autonomía es crucial cuando el estrés o la baja calidad de la grasa comprometen la capacidad digestiva del animal.
En esos momentos críticos, mantener la eficiencia de la digestión lipídica define la rentabilidad del sistema.
LIPA® reduce el tamaño de las gotas de grasa dietética, incrementando sustancialmente la superficie accesible para la lipasa pancreática.
Esto acelera la conversión de triglicéridos en ácidos grasos libres y monoglicéridos, las formas absorbibles del lípido dietético.
Tras la hidrólisis, los ácidos grasos libres y los monoglicéridos requieren integrarse en micelas para su posterior absorción intestinal.
Los LFL estabilizan estas micelas mixtas, optimizando el transporte de los nutrientes lipofílicos a través del lumen.
Al ser más pequeñas y estables, las micelas alcanzan los enterocitos con mayor eficiencia, optimizando la absorción lipídica en el epitelio intestinal (2–4).
Los LFL se integran en la bicapa fosfolipídica de las membranas del enterocito, modulando su fluidez y organización de la membrana.
Esta alteración potencia la absorción de ácidos grasos, nutrientes y vitaminas liposolubles durante el proceso de digestión (5).
En ensayos publicados con pollos de engorde bajo condiciones termoneutras y de estrés calórico, la suplementación con LFL se ha asociado de forma consistente con mejoras:
En la digestibilidad ileal aparente (DIA) de la grasa bruta y la energía bruta.

Esta optimización en el proceso de digestión se traduce en un aumento de peso corporal (APC) y una reducción del índice de conversión alimenticia (ICA) (2,3,6).
Durante la reformulación comercial de la energía dietética, se suele asumir que una reducción en la energía metabolizable inducirá a las aves a incrementar su consumo de alimento como mecanismo de compensación.
Por lo tanto, cualquier déficit energético generado por una menor inclusión de grasa debe recuperarse mediante una mejora en la digestibilidad lipídica, para evitar pérdidas en el rendimiento productivo. En este escenario, los LFL contribuyen a cerrar la brecha energética y proteger los resultados del lote.
Mientras que los lisofosfolípidos (LFL) mejoran la digestión y absorción de los lípidos —sin importar el nivel ni la calidad de la grasa—, estas moléculas dinámicas poseen otras funciones biológicas clave.
Por eso, en la siguiente sección analizamos brevemente su impacto en la integridad de la barrera intestinal, su actividad antioxidante y la salud metabólica del animal.
La integración de los lisofosfolípidos en la membrana del enterocito modifica su organización estructural. Debido a su menor tamaño en comparación con los fosfolípidos estructurales, los LFL generan microespacios dentro de la bicapa, incrementando su fluidez.
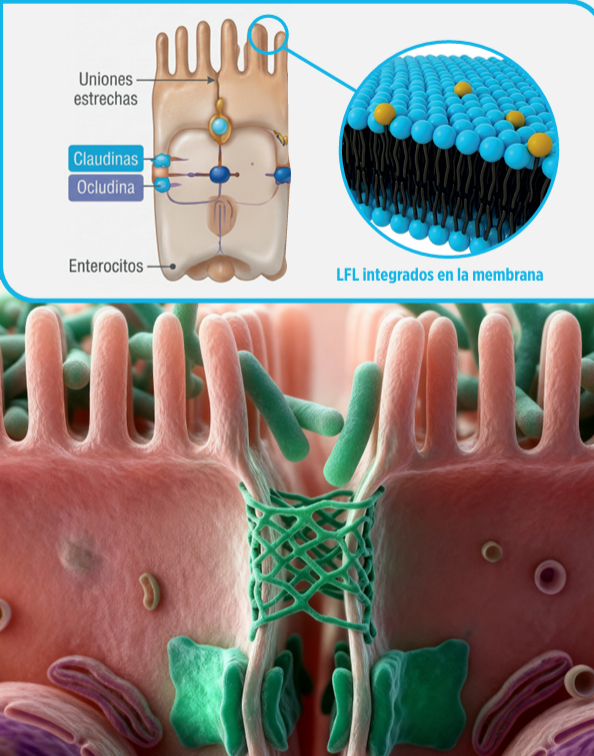
Bajo condiciones de estrés, la expresión de las proteínas de unión estrecha —incluyendo las claudinas y ocludina— disminuye, lo que puede dar lugar a hiperpermeabilidad intestinal, un problema frecuente en la producción aviar (9).
Diversas investigaciones confirman la capacidad de los LFL para revertir las lesiones intestinales.
Inicialmente, Chen et al. (2019) reportaron un incremento en la altura de las vellosidades del intestino delgado y una mayor expresión de Claudina-3 en pollos de engorde suplementados (11).
Posteriormente, Eid et al. (2025) demostraron que, incluso bajo estrés oxidativo, los LFL logran restaurar la arquitectura vellositaria y normalizar los niveles de Claudina-3, vinculando esta mejora con una absorción de nutrientes eficiente y una protección robusta contra patógenos (12).
En definitiva, estos hallazgos posicionan a los LFL como una herramienta útil para apoyar la integridad de la barrera intestinal.
Bajo estrés por altas temperaturas, el exceso de especies reactivas de oxígeno (ERO) supera la capacidad antioxidante del ave, rompiendo el equilibrio celular.
Esta condición provoca la degradación de proteínas y un ataque directo a los lípidos de las membranas celulares.
Durante este proceso de oxidación lipídica, se genera de forma natural el malondialdehído (MDA), un subproducto celular. Al ser liberado al torrente sanguíneo, el MDA se convierte en un biomarcador cuantificable cuya elevación refleja directamente la magnitud del daño tisular y celular (12).
Se ha demostrado que la suplementación con LFL fortalece las defensas antioxidantes del animal.
Este hallazgo fue reportado por Eid et al. (2025) en pollos de engorde sometidos a estrés oxidativo, quienes observaron una actividad significativamente mayor de tres enzimas protectoras: superóxido dismutasa (SOD), catalasa (CAT) y glutatión peroxidasa (GPX).
Además, registraron niveles más bajos de MDA en sangre en comparación con pollos sin suplementación.
El estrés calórico en los pollos de engorde eleva frecuentemente los triglicéridos séricos, el colesterol total y el colesterol de lipoproteínas de baja densidad (LDL-C).
La adición de LFL puede ayudar a contrarrestar este problema, normalizando el perfil lipídico:
Además, en aves de postura y reproductoras, este efecto protector podría ser relevante para reducir el riesgo de síndrome de hígado graso hemorrágico (SGHH), un trastorno metabólico en el que predomina la acumulación excesiva de grasa en el hígado.
El valor de LIPA® se fundamenta en su capacidad para maximizar la digestibilidad de los lípidos. Al optimizar la emulsificación, estabilizar las micelas y favorecer la absorción en la membrana del enterocito, LIPA® recupera eficientemente la energía de la grasa dietética.
Sin embargo, su actividad biológica va mucho más allá del lumen intestinal. Durante el proceso de absorción, los constituyentes de LIPA® se integran estructuralmente en las membranas celulares del enterocito, reforzando las uniones estrechas y consolidando la función de barrera intestinal.
Bajo escenarios de desafío metabólico o ambiental, esta acción potencia las defensas antioxidantes del ave y optimiza su perfil lipídico, reduciendo los factores fisiológicos que pueden desencadenar la inflamación hepática (12,13).
Por lo tanto, el retorno comercial de LIPA® es dinámico y se adapta a las condiciones reales de la granja.
LIPA® no es solo un emulsificante natural: es una herramienta para mantener la consistencia productiva frente a la variabilidad de las materias primas y los desafíos del ambiente. Al favorecer la digestión lipídica, la integridad intestinal y la resiliencia metabólica, ayuda a proteger el retorno económico de la formulación.
Lipa® de Franklin Biotech se comercializa en LATAM bajo el nombre de Gutformance® LIPA a través de nuestro socio comercial ilender. ¡Descubre más aquí! https://avinews.com/franklin-biotech-e-ilender-se-unen-para-impulsar-la-salud-intestinal/
Materias Primas


Suscribete ahora a la revista técnica de avicultura
AUTORES

Empresas GEEK – Pautas aplicables en la Prefaena y el Procesamiento para aumentar la Productividad
Eduardo Cervantes López
¿Qué puede aprender LATAM de la adopción de tecnologías de sexado en Europa?
Equipo Técnico TARGAN
El Eje Intestino-Hígado: El Motor de la Eficiencia Metabólica y Bioseguridad Interna en Avicultura
Joan Rodríguez MV
Optimizando la nutrición del pollo de engorde moderno – Parte II
Xavier Asensio
Versatilidad del tamaño del huevo en NICK CHICK – Parte I
Equipo Técnico H&N
Valor agregado en la industria avícola: El potencial del pollo en embutidos cocidos
Equipo Avinews Latam
Estrategias de Bioseguridad Integrales para Control de la Influenza Aviar de Alta Patogenicidad: Vacunación Estratégica y Monitoreo Zoonótico en Aves Silvestres II
MVZ. Cesar Isaac Ascencio Pérez MVZ. José Alejandro Jiménez Ramírez
Plumas, Ventiladores y Temperatura ambiental: ¡Guía práctica para el confort de los pollitos!
Udaykumar (Uday Patil) Mudbakhe
Longitud de las vellosidades intestinales del yeyuno e íleon en pollos de engorde
Dra. Odette Urquiza Bravo Félix Sánchez Godoy Guillermo García Hernández Nestor Ledesma Martínez
Recubrimientos sostenibles para reducir las pérdidas de huevos
Gabriel da S. Oliveira Paula Gabriela da Silva Pires Priscila de O. Moraes Vinicius M. dos Santos
Implementación de formulación dinámica de alimentos balanceados y nutrición de precisión en la producción avícola
Edgar O. Oviedo Rondón
Latidos programados: Cómo la temperatura y la hipoxia durante la incubación moldean el corazón y el sistema vascular del pollito
Cristiano Emanuelli Pereira Dr. Marcelo Leite da Veiga Isabela Maraschin Vieira
True Performance: El fin de los altibajos en el rendimiento avícola
Equipo Técnico BAADER






































